Play all audios:
ABSTRACT We recently identified genes and molecular pathways related to radioresistance of oral squamous cell carcinoma (OSCC) using Affymetrix GeneChip. The current study focused on the
association between one of the target genes, intercellular adhesion molecule 2 (_ICAM2_), and resistance to X-ray irradiation in OSCC cells, and evaluated the antitumor efficacy of combining
ICAM2 small interfering RNA (siRNA) and X-ray irradiation. Downregulation of ICAM2 expression by siRNA enhanced radiosensitivity of OSCC cells with the increased apoptotic phenotype via
phosphorylation (ser473) of AKT and activation of caspase-3. Moreover, overexpression of ICAM2 induced greater OSCC cell resistance to the X-ray irradiation with the radioresistance
phenotype. These results suggested that ICAM2 silencing is closely related to sensitivity of OSCC cells to radiotherapy, and that ICAM2 may be an effective radiotherapeutic target for this
disease. SIMILAR CONTENT BEING VIEWED BY OTHERS SALL4 PROMOTES CANCER STEM-LIKE CELL PHENOTYPE AND RADIORESISTANCE IN ORAL SQUAMOUS CELL CARCINOMAS VIA METHYLTRANSFERASE-LIKE 3-MEDIATED M6A
MODIFICATION Article Open access 14 February 2024 YY1 MODULATES THE RADIOSENSITIVITY OF ESOPHAGEAL SQUAMOUS CELL CARCINOMA THROUGH KIF3B-MEDIATED HIPPO SIGNALING PATHWAY Article Open access
08 December 2023 B7-H3 REGULATES KIF15-ACTIVATED ERK1/2 PATHWAY AND CONTRIBUTES TO RADIORESISTANCE IN COLORECTAL CANCER Article Open access 03 October 2020 MAIN Radiation therapy has played
an important role in controlling tumour growth in many patients with cancer. In patients with oral squamous cell carcinoma (OSCC), radiation therapy is currently the standard adjuvant
treatment. However, radiation therapy is sometimes ineffective as cancer cells can be resistant to radiation. Our previous study showed that the radiosensitivity of OSCC cell lines differs
greatly in their response to X-ray radiation, as assessed by clonogenic survival assay (Higo et al, 2005). In this study, we showed that in six OSCC cell lines HSC2 was the most
radioresistant and HSC3 was the most radiosensitive cell line. Furthermore, we showed that 25 genes and their related molecular pathways related to cell growth and antiapoptosis were
strongly associated with radiation resistance in OSCC (Ishigami et al, 2007). Of them, six were reported to be related to cell proliferation and antiapoptosis, that is, _ID1_ (Nakajima et
al, 1998; Zhang et al, 2004b), _ID3_ (Plowright et al, 2000; Kowanetz et al, 2004), _FGFR3_ (Gómez-Román et al, 2005), _PEG10_ (Okabe et al, 2003; Tsou et al, 2003; Hu et al, 2004),
intercellular adhesion molecule 2 (_ICAM2_) (Perez et al, 2002), and _MMP13_ (McCawley and Matrisian, 2000; Chang and Werb, 2001; Corte et al, 2005), and we validated that the expression
levels increased substantially in radioresistant cells compared with radiosensitive cells. Among the six genes identified, _ICAM2_, whose mRNA level in unirradiated HSC2 (radioresistant
cells compared with unirradiated HSC3 (radiosensitive) cells was highest in our previous microarray results (Ishigami et al, 2007), underwent further functional analysis. Intercellular
adhesion molecule 2 has been thought to play a role in lymphocyte recirculation by blocking LFA-1-dependent cell adhesion and mediating adhesive interactions important for an
antigen-specific immune response, NK cell-mediated clearance, lymphocyte recirculation, and other cellular interactions important for immune response and surveillance (Staunton et al, 1989;
Li et al, 1995; Helander et al, 1996; Carpenito et al, 1997; Lehmann et al, 2003). However, ICAM2 has been reported to be a mediator for a survival signal sufficient to block apoptosis by
activation of the PI3K/AKT pathway (Perez et al, 2002). In addition, another recent study has reported that absence of ICAM2 expression resulted in impaired angiogenesis _in vitro_ and _in
vivo_, and that ICAM2-deficient cells were defective in _in vitro_ migration and increased apoptosis (Huang et al, 2005). It has been shown that _ICAM2_-expressing cells may be resistant to
apoptosis induced by anticancer agents, including radiation. Therefore, we hypothesised that _ICAM2_ inhibitors could enhance the effect of radiation on cancer cells that constitutively
express _ICAM2_. Small-interfering RNA has been used widely to silence gene expression and has been evaluated as an attractive tool for use in therapeutics of many cancers (Gao et al, 2005;
Zhang et al, 2005; Amarzguioui et al, 2006; Halder et al, 2006; Hosaka et al, 2006). So, we evaluated whether radiosensitivity of oral cancer cells is determined by cellular _ICAM2_ by using
siRNA targeted against _ICAM2_ gene. Furthermore, we analysed the radiosensitivity of OSCC cells by upregulating _ICAM2_ gene expression using expression vector encoding ICAM2 cDNA.
MATERIALS AND METHODS CELL LINES AND CULTURE CONDITIONS The human OSCC-derived cell lines HSC2 and HSC3 (Human Science Research Resources Bank, Osaka, Japan) were prepared for this study.
The cells were maintained in Dulbecco's modified Eagle's medium (DMEM) (Sigma Chemical Co., St Louis, MO, USA) supplemented with 10% heat-inactivated foetal bovine serum and 50 U
ml−1 penicillin and streptomycin. All cultures were grown at 37°C under a humidified atmosphere of 5% carbon dioxide for routine growth. SIRNA, TRANSFECTION REAGENTS, AND TRANSFECTION OF
SIRNAS IN HSC2 (RADIORESISTANT) CELLS Small-interfering RNAs were obtained from Dharmacon Research Inc. (Lafayette, CO, USA). _SMART_ pool siRNA targeting _ICAM2_ consists of four siRNAs
targeting multiple sites on _ICAM2_ (_ICAM2_-siRNAs). The sequences for _ICAM2_-siRNAs are 5′-AAGCAGGAGUCAAUGAAUU-3′, 5′-UAACCAGCCUGAAGUGGGU-3′, 5′-UGAGAAGGUAUUCGAGGUA-3′, and
5′-ACGAACAGGCUCAGUGGAA-3′ (si_GENOME SMART_pool, M-11182-00-0005, Human _ICAM2_, NM_000873). Positive- and negative-control siRNAs were purchased from Dharmacon. Two negative controls were
used in this study, that is, vehicle control (treated with only Dharma_FECT_1 reagent) and si_CONTROL_ non-targeting siRNA pool (D-001210-01-05; non-targeting siRNA (siNT)). Cyclophilin _β_
(siCONTROL Cyclophilin _β_ siRNA, D-001136-01-05) was used as positive silencing control to ascertain transfection efficiency. Cells were transfected with siRNAs using Dharma_FECT_1 reagent
(Dharmacon). To confirm whether _ICAM2_ gene is related to radioresistance, we performed an siRNA experiment to inhibit the expression of _ICAM2_ in HSC2 (radioresistant) cell line that
previously reported the expression of _ICAM2_ as being higher than HSC3 (radiosensitive) cell line (Ishigami et al, 2007). HSC2 Cells were plated in antibiotic-free DMEM at a density of 200
000 cells 4 ml−1 in 60-mm dishes. After 24 h, the cells were transfected with 100 nmol l−1 siRNA in Dharma_FECT_1 reagent according to the manufacturer's instructions. Briefly, 8 _μ_l
Dharma_FECT_1 was diluted in 392 _μ_l of serum-free medium and incubated at room temperature for 5 min. In a separate tube, 200 _μ_l of 2 _μ_mol l−1 siRNA was diluted in 200 _μ_l of
serum-free medium at room temperature for 5 min. Diluted Dharma_FECT_1 (400 _μ_l) was added to the diluted siRNA and the complex was incubated for 20 min at room temperature. The cells were
washed with antibiotic-free DMEM and 3.2 ml antibiotic-free DMEM was added to each dish. Small interfering RNA+Dharma_FECT_1 complex (800 _μ_l) was added gently to the dish. The final
concentration of siRNA was 100 nmol l−1. Control cells were treated with the only medium (HSC2 control), the same amount of transfection reagents (vehicle control), the 100 nmol l−1 siNT
(negative control), and the 100 nmol l−1 cyclophilin _β_ siRNA (siCyclophilin _β_) (positive silencing control). After 4 h of transfection, the medium of cells treated with _ICAM2_-siRNAs
(siRNA targeted to ICAM2 (siICAM2)) and control cells was replaced with fresh medium, and these were incubated at 37°C in 5% CO2 for 48 h to 120 h before performing experiments. TRANSIENT
TRANSFECTION OF ICAM2 DNA To verify whether _ICAM2_ gene is related to radioresistance, we performed overexpression of _ICAM2_ gene in HSC3 (radiosensitive) cell line that previously
reported the expression of _ICAM2_ as being higher than HSC2 (radioresistant) cell line (Ishigami et al, 2007). Human _ICAM2_ cDNA was cloned into a pME18SFL3 expression vector (TOYOBO,
Osaka, Japan) for transient transfection experiments. HSC3 cell lines were transfected with pME18SFL3 encoding _ICAM2_ cDNA using the FuGENE HD transfection reagent (Roche Diagnostics GmbH,
Mannheim, Germany). Mock transfection of HSC3 cell line cultures with the FuGENE HD transfection reagent alone was used as vehicle controls. Transfection efficiency was confirmed by
real-time quantitative reverse transcriptase–polymerase chain reaction (qRT–PCR) and western blot analysis. These analyses were performed as described below. IRRADIATION USING X-RAY The
cells were irradiated with four single radiation doses (2, 4, 6, and 8 Gy) using X-ray irradiation equipment (MBR-1520R-3; Hitachi, Tokyo, Japan) operated at 150 V and 20 mA with AL
filtration, at a dose of 2.1 Gy min−1. ISOLATION OF RNA Total RNA was extracted from X-ray-irradiated and unirradiated cells with TRIzor reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA) according to the manufacturer's instructions. The quality of the total RNA was determined using Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). PREPARATION OF CDNA Total
RNA was extracted from cells using TRIzor reagent. Five micrograms of total RNA of each sample were reversed transcribed to cDNA using Ready-To-Go You-Prime First-Strand Beads (GE
Healthcare, Little Chalfort, Buckinghamshire, UK) and oligo (dT) primer (Sigma Genosys, Ishikari, Japan), according to the manufacturers’ protocol. ANALYSIS OF MRNA EXPRESSION BY REAL-TIME
QRT–PCR Quantitative reverse transcriptase–polymerase chain reaction was performed to validate mRNA expression with a single method using a LightCycler FastStart DNA Master SYBR Green I kit
(Roche Diagnostics GmbH), according to the procedure provided by the manufacturer. The oligonucleotides used as primers were 5′-GATCCAGGGCGGAGACTTC-3′ and 5′-GCCCGTAGTGCTTCAGTTTGA-3′ for
_Cyclophilin β_ mRNA, 5′-CATCTCTGCCCCCTCTGCTGA-3′ and 5′-GGATGACCTTGCCCACAGCCT-3′ for glyceraldehyde-3-phosphate dehydrogenase (_GAPDH_) mRNA, and 5′-ATTCAACAGCACGGCTGACA-3′ and
5′-CAGGCTCATAGATCTCCAACATCT- 3′ for _ICAM2_ mRNA. Using LightCycler apparatus, we carried out PCR reactions in a final volume of 20 _μ_l of a reaction mixture consisting of 2 _μ_l of
FastStart DNA Master SYBR Green I mix, 3 mM MgCl2, and 1 _μ_l of the primers, according to the manufacturer's instructions. Subsequently, the reaction mixture was loaded into glass
capillary tubes and subjected to initial denaturation at 95°C for 10 min, followed by 33–45 rounds of amplification at 95°C (10 s) for denaturation, 62–68°C (10 s) for annealing, and 72°C
for extension, with a temperature slope of 20°C s−1, performed with LightCycler. The transcript amount for the genes was estimated from the respective standard curves and normalised to the
_GAPDH_ transcript amount determined in the corresponding samples. PROTEIN EXTRACTION Protein was extracted from the cells, which were washed twice with phosphate-buffered saline, scraped
into a tube with lysis buffer (7 M urea, 2 M thiourea, 4% w/v CHAPS, and 10 mM Tris, pH 8), and incubated at 4°C for 10 min. Cell extracts were lysed by sonication (3 × 10-s pulses on ice)
and centrifuged at 13 000 G for 10 min at 4°C. The supernatant containing the cell proteins then was recovered. Protein concentration was determined using a commercial Bradford reagent
(Bio-Rad, Richmond, CA, USA) and adjusted to 1 mg ml−1 with lysis buffer. WESTERN BLOT ANALYSIS Protein extracts (15 _μ_g) were electrophoresed on 12.5% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis gels, transferred to polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA, USA), and blocked for 1 h at room temperature in 0.3% skimmed milk. Immunoblot
PVDF membranes were washed with Tris-buffered saline Tween-20 (TBST: 10 mM Tris-HCl (pH 8.5), 150 mM NaCl, and 0.1% Tween-20) thrice and probed with 2 _μ_g ml−1 affinity-purified mouse
anti-human ICAM2 monoclonal antibody (R&D Systems Inc., Minneapolis, MN, USA), 1 _μ_g ml−1 affinity-purified rabbit anti-human/mouse/rat specific AKT antibody (Rockland Inc.,
Gilbertsville, PA, USA), and 1 _μ_g ml−1 affinity-purified mouse anti-human p-AKT (pS437) antibody (BIOMOL International, L.P., Plymouth Meeting, PA, USA) overnight at room temperature. For
cyclophilin _β_ protein and _β_-actin protein, 1 _μ_g ml−1 affinity-purified rabbit anti-human cyclophilin _β_ polyclonal antibody (Abcam Ltd, Cambridge, UK) and 1 _μ_g ml−1
affinity-purified mouse anti-human _β_-actin monoclonal antibody (Abcam Ltd) were used for 2 h at room temperature. Polyvinylidene difluoride membranes were washed again and incubated with 1
: 10 000 of Envision+(Dako Japan Inc., Kyoto, Japan) as secondary antibody coupled to horseradish peroxidase-conjugated anti-mouse or ant-rabbit IgG antibody for 2 h at room temperature.
Finally, membranes were incubated using the ECL Plus Western Blotting Detection System (GE Healthcare, Carpinteria, Buckinghamshire, UK) and immunoblotting result was visualised by exposing
the membrane to Fuji Medica X-Ray film RX-U (FUJIFILM, Kanagawa, Japan). Signal intensities were quantitated using ImageJ software (1.38x version)(NIH, Bethesda, MD, USA). CELL-PROLIFERATION
ASSAY To determine the effect of _ICAM2_ siRNA on cell proliferation, HSC2 cells transfected with non-targeting or _ICAM2_ siRNA (100 nmol l−1) were seeded in 12-well plates at a density of
1 × 104 viable cells per well. Mock-transfected cells were treated with Dharma_FECT_1 reagent as vehicle controls but not siRNA. At the indicated time point, cells were trypsinised and
counted using a haemocytometer in triplicate samples. The effect on cell proliferation was investigated in the same manner as HSC3 cells treated with expression vector of _ICAM2_ DNA.
CLONOGENIC CELL SURVIVAL ASSAY HSC2 cells were transfected as above with the vehicle, siNT, and siICAM2. At 72, 84, and 96 h after transfection, the cells were trypsinised, counted, and the
appropriate number of cells were plated in 60-mm dishes and allowed to attach for 24 h. After 24 h, the cells were irradiated (2, 4, 6, 8 Gy) and incubated for 8–10 days. The colonies were
stained with crystal violet (Sigma Chemical Co.), and colonies of 50 cells or greater were counted. Clonogenic fractions of irradiated cells were normalised to the plating efficiency of
unirradiated controls. Likewise, colonies of HSC3 cells transfected with expression vector of _ICAM2_ DNA were measured. _IN SITU_ CELL APOPTOSIS DETECTION ASSAY HSC2 cells were plated in
24-well dishes at a density of 2 × 104 cells per well. After 24 h, HSC2 cells were treated with siRNA (vehicle, siNT, siICAM2). After incubation for 120 h, cells were analysed for apoptosis
by terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick-end labelling (TUNEL) procedure, using an _in situ_ apoptosis detection kit (Takara Bio, Shiga, Japan). Briefly, cells were
fixed in 4% paraformaldehyde for 15 min. After treatment with permeabilisation buffer (Takara Bio) for 2 min on ice, the cells were incubated with TdT enzymes plus labeling safe buffer
(Takara Bio) at 37°C. After 1.5 h, the cells were washed with phosphate-buffered saline and observed by fluorescence microscopy. Cells undergoing apoptosis were specifically labelled with
fluorescein-dUTP with high sensitivity, allowing immediate detection by fluorescence microscopy. Apoptotic cells were counted and expressed as a percentage of the total cells. The
experiments were performed in triplicate. CASPASE-3 ACTIVITY ASSAY We performed three different assays for caspase-3 activity. Two assays (using caspase-3 detection kit; Calbiochem, Oxford,
UK) measured the amount of activated caspase-3 _in situ_ in living cells by a caspase-3 inhibitor (DEVD-FMK) conjugated to FITC as a fluorescent _in situ_ marker. Fluorescein
isothiocyanate-DEVD-FMK is cell-permeable and nontoxic and irreversibly binds to activated caspase-3 in apoptotic cells. The FITC label allows direct detection of activated caspase-3 by
fluorescence microscopy or a fluorescence microplate reader. In our initial experiment, FITC fluorescence was conjugated to the inhibitor, thereby registering a fluorescent label detected by
fluorescence microscopy in cells in which caspase-3 had been activated. Both cells treated with and without ICAM2 siRNA (control, vehicle, and siNT) were compared. Caspase-3 active cells
were counted and expressed as a percentage of the total number of cells. The experiments were performed in triplicate. For the next experiment, we used a microplate reader to study the
effect. Data were expressed as the difference in activity among the four data points (control, vehicle, siNT, and siICAM2) and indicated the amount of caspase-3 activity that remained. The
kit was used according to the manufacturer's instructions. Briefly, cells were treated with or without ICAM2 siRNA for 120 h, after which they (about 1 × 106 ml−1) were incubated for 1
h with FITC-DEVD-FMK at 37°C. The cells then were washed twice by washing buffer and then the fluorescence intensity was measured by fluorescence microscopy and using a fluorescence
microplate reader (Wallac 1420 ARVOsx Multilabel Counter; Perkin-Elmer, Chiba, Japan), with excitation at approximately 485 nm and emission detection at approximately 535 nm. Finally, we
performed activated caspase-3 western blot analysis. Monoclonal antibody to active caspase-3 (Genlantis Inc., San Diego, CA, USA) was used for detection. Western blot analysis for activated
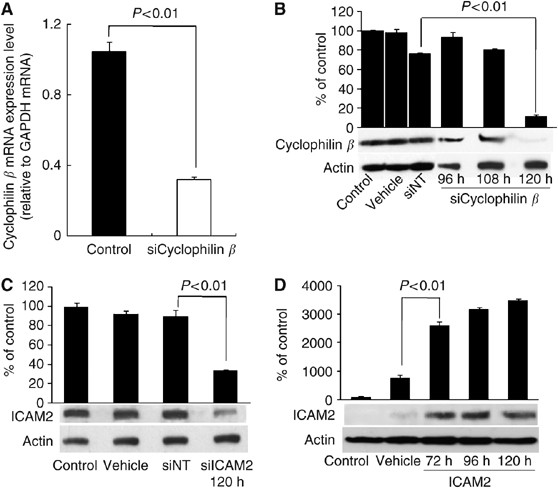
caspase-3 was performed as described previously. RESULTS TRANSFECTION EFFICIENCY OF SIRNAS To ascertain whether conditions for RNA inhibition were optimal and that transfection efficiency
was satisfactory, siCyclophilin _β_ was used as a positive silencing control. In real-time qRT–PCR analysis, _cyclophilin β_ mRNA expression decreased by 69% with siCyclophilin _β_ 48 h
after transfection (Figure 1A). In western blot analysis, cyclophilin _β_ was detected as a single band. In cells transfected with siCyclophilin _β_, the band diminished significantly
(_P_<0.01, Student's _t_-test), confirming high transfection efficiency 120 h after transfection compared with the vehicle, si_CONTROL_ siNT, and the non-transfected control cells.
HSC2 cells transfected with siCyclophilin _β_ showed little change in the band of cyclophilin _β_ 96 and 108 h after transfection compared with controls (Figure 1B). ICAM2 PROTEIN LEVELS IN
HSC2 CELLS TRANSFECTED WITH SIICAM2 AND CONTROL SIRNAS Figure 1C shows western blot analysis of ICAM2 protein expression in HSC2 cells 120 h after transfection with siRNAs. This time point
was selected by a positive silencing control test in an siRNA transfection experiment (Figure 1B). The ICAM2 protein levels in cells transfected with vehicle, siNT, and siICAM2-transfected
cells were comparable to those in non-transfected cells. In cells transfected with 100 nmol l−1 siICAM2, ICAM2 protein level decreased significantly (_P_<0.01, Student's _t_-test)
compared with non-transfected, vehicle-transfected, and the siNT-transfected cells (Figure 1C). In addition, qRT–PCR analysis showed that _ICAM2_ mRNA expression decreased by 67, 97, and 96%
in HSC2 cells with siICAM2 compared with non-transfected cells at 24, 48, and 72 h, respectively, after transfection. ICAM2 PROTEIN LEVELS IN HSC3 CELLS TRANSFECTED WITH ICAM2 DNA Figure 1D
shows western blot analysis of ICAM2 protein expression in HSC3 cells 72, 96, and 120 h after transfection with expression vector encoding _ICAM2_ cDNA. The ICAM2 protein level increased
significantly (_P_<0.01, Student's _t_-test) in cells transfected with _ICAM2_ DNA compared with vehicle and non-transfected control cells (Figure 1D). Furthermore, qRT–PCR analysis
showed that the level of _ICAM2_ mRNA expression increased significantly (_P_<0.01, Student's _t_-test) by 4.5 × 106 and 9.4 × 106-fold in HSC3 cells with expression vector encoding
ICAM2 cDNA compared with non-transfected cells 48 and 72 h, respectively, after transfection. EFFECT OF ICAM2 SIRNA AND EXPRESSION VECTOR ENCODING ICAM2 CDNA ON CELL GROWTH To determine the
effect of _ICAM2_ siRNA and expression vector encoding _ICAM2_ cDNA on the growth of the cancer cell lines, a series of cell growth experiments was carried out. The growth curves of HSC2
cells showed that treatment with ICAM2 siRNA inhibited cell growth over a period of 6 days, but cell growth was not inhibited by vehicle and siNT (Figure 2A). Furthermore, the growth curve
of HSC3 cells showed that treatment with expression vector encoding _ICAM2_ cDNA increased cell growth over a period of 6 days compared with the vehicle (Figure 2B). RADIOSENSITIVITY OF HSC2
CELLS TRANSFECTED WITH SIICAM2 The plating efficiencies of unirradiated HSC2 cells (mean±s.d., _n_=3) were 0.278±0.038 (control), 0.270±0.032 (vehicle), 0.262±0.031 (siNT), 0.258±0.031
(siICAM2, 120 h), 0.275±0.066 (siICAM2, 108 h), and 0.220±0.057 (siICAM2, 96 h). The difference in the plating efficiency of siICAM2 cells compared with vehicle or siNT cells was not
significant. Figure 2C shows the radiation survival curves of untreated control cells and cells transfected with siRNAs. These data were used to calculate dose D37 that is required to reduce
survival to 37% (Toulany et al, 2006). For HSC2 cells, dose D37 values for control, vehicle, siNT, siICAM2 at 108 h, siICAM2 at 96 h, and siICAM2 at 120 h were 7.6, 7.6, 7.7, 6.0, 4.7, and
4.4 Gy, respectively. Compared with the vehicle-treated control, at the 37% survival level, the radiosensitivity of cells transfected with siICAM2 at 120 h was enhanced by a dose-modifying
factor of 1.73 and the radiosensitivity of cells transfected with siNT was 0.99-fold. CELLS TRANSFECTED WITH SIICAM2 AT 120 H COMPARED WITH CELLS TRANSFECTED WITH SINT The radiosensitivity
of cells transfected with siICAM2 at 120 h was enhanced by a dose-modifying factor of 1.75 for siNT. Moreover, survival of HSC2 cells transfected with siICAM2 at 120 h decreased
significantly (_P_<0.01, Student's _t_-test) after 4, 6, and 8 Gy of radiation compared with that of HSC2 cells treated with siNT (Figure 2C). This indicates that radiation-induced
sublethal damage is severe when _ICAM2_ expression is inhibited. RADIOSENSITIVITY OF HSC3 CELLS TRANSFECTED WITH EXPRESSION VECTOR OF ICAM2 DNA In HSC3 cells, the plating efficiencies at 0
Gy (mean±s.d., _n_=3) were 0.211±0.030 (control), 0.262±0.003 (vehicle), and 0.232±0.030 (ICAM2). The difference in the plating efficiency of cells transfected with expression vector of
_ICAM2_ DNA, compared with vehicle or untreated control cells, was not significant. The radiation survival curves of cells are shown in Figure 2D. In cells transfected with expression vector
encoding ICAM2 cDNA, compared with cells transfected with vehicle and untreated control cells, the radiosensitivity of the transfectant decreased. For HSC3 cells, dose D37 values for
control, vehicle, and the transfectant were 3.4, 4, and 5 Gy, respectively. The resulting dose-modifying factors were 0.80 for the vehicle and 0.68 for the untreated control cells at the 37%
survival level. Furthermore, the rate of cell survival of the transfectant increased significantly (**_P_<0.01, *_P_<0.05, Student's _t_-test) after 2, 4, 6, and 8 Gy of
radiation, compared with that of cells treated with vehicle (Figure 2D). CORRELATION OF ICAM2 EXPRESSION AND APOPTOSIS To examine whether expression of _ICAM2_ is the result of apoptosis, we
analysed DNA fragmentation by TUNEL assay. As shown in Figure 3A–D, positive TUNEL labelling in apoptotic cells was bright. Figure 3D clearly shows that apoptosis was induced in some cells
but not in others (Figure 3A–C). The percentages of apoptotic cells in HSC2 cells transfected with the vehicle, siNT, siICAM2, and in the untreated control cells were 1.40±0.09, 2.56±0.29,
26.4±8.81, and 1.05±0.14%, respectively. Data are expressed as the mean±s.d. of three independent experiments. The difference between siNT and siICAM2 reached significance (_P_<0.01,
Student's _t_-test). ICAM2-INDUCED AKT PHOSPHORYLATION Recent studies have reported that ICAM2 activated the protein kinase B (PKB)/AKT pathway leading to inhibition of apoptosis (Perez
et al, 2002). In this pathway, phosphorylation (Ser473) of AKT protein induced antiapoptosis. To investigate the correlation of ICAM2 and AKT phosphorylation, we performed western blot
analysis with antibody recognising AKT and AKT phosphorylation at Ser473 site (Figure 3I). Figure 3I shows that HSC2 cells treated with siICAM2 resulted in lower AKT phosphorylation level
than cells treated with the vehicle, siNT, and the untreated control cells. CASPASE-3 ACTIVITY ASSAY We investigated the role of caspase-3 in this process to determine the mechanism by which
apoptosis occurred. Figure 4A–D show the green FITC signal present within the siICAM2 cells; control, vehicle, and siNT cells displayed a small activated caspase-3 signal. The percentages
of caspase-3 active cells in HSC2 cells transfected with the vehicle, siNT, siICAM2, and the untreated control cells were 2.60±2.30%, 4.00±0.61%, 20.7±0.71%, and 1.45±2.51%, respectively.
Data are expressed as the mean±s.d. of three independent experiments. The difference between siNT and siICAM2 reached significance (_P_<0.01, Student's _t_-test). The second assay
(caspase-3 activity assay) (Figure 4I) showed that caspase-3 activity was significantly elevated in cells with ICAM2 siRNA for 120 h compared with cells with vehicle and siNT (_P_<0.05,
Student's _t_-test). Quantification of relative fluorescence intensity revealed that siICAM2 increased caspase-3 enzymatic activity 1.36-fold compared with siNT. The third study
examined the protein levels of active caspase-3. Figure 4J shows that HSC2 cells treated with siICAM2 had significantly higher (_P_<0.01, Student's _t_-test) active caspase-3 protein
levels than cells treated with the vehicle, siNT, and the untreated control cells. DISCUSSION _ICAM2_, whose mRNA level was upregulated in radioresistant OSCC cells in our previous study
(Ishigami et al, 2007), was one of candidate genes selected as radioresistant genes of OSCC cells by microarray analysis using Affymetrix GeneChip. The current study was designed to examine
whether ICAM2 is functionally associated with radiosensitivity of OSCC _in vitro_. In the previous report, little has been mentioned about the role of ICAMs in regard to radioresistance.
Intercellular adhesion molecule 1 expression was shown to be elevated by hypoxia and radiation (Zünd et al, 1997; Meineke et al, 2002). Intercellular adhesion molecule 3 expression was
reported to associate with radioresistance in cervical cancer (Chung et al, 2005). In our study, the results indicated that ICAM2 inhibition induces radiosensitive _in vitro_ (Figure 2C). A
further important point is that radiosensitisation of cells may depend on the extent of ICAM2 protein inhibition. Because HSC2 cells were incubated for a longer time with siICAM2, they
became more radiosensitive (Figure 2C), suggesting that radiosensitisation of HSC2 cells may be determined by the amount of ICAM2 protein present in the cells at the time of irradiation.
Moreover, ICAM2 overexpression induces radioresistance _in vitro_ (Figure 2D). These results may indicate that regulation of ICAM2 protein is correlated with radiosensitivity. Further,
Figure 2A and B show that regulation of ICAM2 protein is related to cell growth. The question that needs to be answered is what mechanisms play a role in the relation between the regulation
of the ICAM2 protein and these results. Intercellular adhesion molecule 2-induced activation of AKT kinase resulted in the activation of several downstream effectors as detected by
phosphorylation of BAD, GSK3, FKHR, and AFX, all of which can contribute to cell survival (Zha et al, 1996; Brunet et al, 1999). The PI3/AKT-signalling system is a general mediator of
extracellular stimuli, including growth factors, cytokines, and adhesion, to extracellular matrices (Downward, 1998). Phosphatidylinositol-3,4,5-triphosphate binds to the pleckstrin domain
of AKT and recruits AKT to the membrane, where it becomes dually phosphorylated (Alessi et al, 1996) and is then activated and phosphorylates a number of downstream effectors that contribute
to cell survival. These studies indicated that ICAM2 activation might lead to an antiapoptic signal in a variety of cells. A recent study identified a pathway of ICAM2 activating the
PI3K/AKT leading to inhibition of apoptosis (Perez et al, 2002). This pathway showed that ICAM2 induced tyrosine phosphorylation of ezrin and PI3K kinase membrane translocation, resulting in
phosphatidylinositol-3,4,5 production, PDK-1 and AKT activation, and subsequent phosphorylation of AKT targets BAD, GSK3, and FKHR. The previous studies may have presumed that the
differences in cell growth and radiosensitivity are caused by mechanisms of antiapoptotic effect induced by ICAM2. Consequently, we postulated that ICAM2 induced antiapoptosis in OSCCs as
well as the mechanism of the PKB/AKT pathway activation previously reported (Perez et al, 2002). In addition, AKT phosphorylation at Ser473 was suggested to be correlated perfectly with
antiapoptosis in lymphocytes (Perez et al, 2002). Therefore, we examined AKT phosphorylation (Ser473) and cell apoptosis to confirm the relation between ICAM2 and antiapoptosis via the
PKB/AKT pathway in OSCC cells. The results of our experiment showed that cells with siICAM2 induced a decrease in AKT phosphorylation (Ser473) (Figure 3I). To detect cell apoptosis, we
performed caspase-3 activity assays and TUNEL assay. We evaluated caspase-3 activation in cells treated with ICAM2 siRNA. HSC2 cells treated with siICAM2 showed increased caspase-3 activity
compared with untreated or control cells (Figure 4). In the previous studies, the apoptotic pathway induced by decrease in p-AKT caused activation of caspase-3 (Ivins Zito et al, 2004; Zhang
et al, 2004a; Zhuang et al, 2007). We considered that decreased p-AKT and increased activated caspase-3 induced apoptosis in cells treated with ICAM2 siRNA. Furthermore, results of the
TUNEL assay showed increased apoptosis (Figure 3D). Accordingly, it is reasonable that inhibiting ICAM2 may contribute to radiosensitisation of OSCC cells by increased apoptosis via
phosphorylation (Ser473) of AKT and activation of caspase-3. Our study was similar to a previous study (Chung et al, 2005) in which the investigators used the same methods, that is,
microarray analysis, siRNA, overexpression, and apoptosis assay. These two studies yielded similar results in that they both confirmed a relation between a gene and radioresistance. However,
due to availability of clinical samples, our study differed from the other in that we could not analyse clinical specimens from patients. Our study, therefore, may be insufficient _in
vivo_. Our study could not prove a relation between ICAM2 and radioresistance in OSCC cells _in vitro_. In conclusion, _ICAM2_ expression may be associated with radioresistance in OSCC
cells, _ICAM2_ siRNA may enhance the radiosensitivity of oral cancer cells, and ICAM2 may be an effective radiotherapeutic target of oral cancer and a marker for radiation sensitivity based
on _in vitro_ studies with microarray analysis. CHANGE HISTORY * _ 16 NOVEMBER 2011 This paper was modified 12 months after initial publication to switch to Creative Commons licence terms,
as noted at publication _ REFERENCES * Alessi D, Andjelkovic M, Caudwell B, Cron P, Morrice N, Cohen P, Hemmings B (1996) Mechanism of activation of protein kinase B by insulin and IGF-1.
_EMBO J_ 15: 6541–6551 Article CAS PubMed PubMed Central Google Scholar * Amarzguioui M, Peng Q, Wiiger M, Vasovic V, Babaie E, Holen T, Nesland J, Prydz H (2006) _Ex vivo_ and _in
vivo_ delivery of anti-tissue factor short interfering RNA inhibits mouse pulmonary metastasis of B16 melanoma cells. _Clin Cancer Res_ 12: 4055–4061 Article CAS PubMed Google Scholar *
Brunet A, Bonni A, Zigmond M, Lin M, Juo P, Hu L, Anderson M, Arden K, Blenis J, Greenberg M (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription
factor. _Cell_ 96: 857–868 Article CAS PubMed Google Scholar * Carpenito C, Pyszniak A, Takei F (1997) ICAM-2 provides a costimulatory signal for T cell stimulation by allogeneic class
II MHC. _Scand J Immunol_ 45: 248–254 Article CAS PubMed Google Scholar * Chang C, Werb Z (2001) The many faces of metalloproteases: cell growth, invasion, angiogenesis and metastasis.
_Trends Cell Biol_ 11: S37–S43 Article CAS PubMed PubMed Central Google Scholar * Chung Y, Kim B, Park C, Huh S, Kim J, Park J, Cho S, Kim B, Kim J, Yoo Y, Bae D (2005) Increased
expression of ICAM-3 is associated with radiation resistance in cervical cancer. _Int J Cancer_ 117: 194–201 Article CAS PubMed Google Scholar * Corte M, Gonzalez L, Corte M, Quintela I,
Pidal I, Bongera M, Vizoso F (2005) Collagenase-3 (MMP-13) expression in cutaneous malignant melanoma. _Int J Biol Markers_ 20: 242–248 Article CAS PubMed Google Scholar * Downward J
(1998) Mechanisms and consequences of activation of protein kinase B/Akt. _Curr Opin Cell Biol_ 10: 262–267 Article CAS PubMed Google Scholar * Gao L, Zhang L, Hu J, Li F, Shao Y, Zhao
D, Kalvakolanu D, Kopecko D, Zhao X, Xu D (2005) Downregulation of signal transducer and activator of transcription 3 expression using vector-based small interfering RNAs suppresses growth
of human prostate tumor _in vivo_. _Clin Cancer Res_ 11: 6333–6341 Article CAS PubMed Google Scholar * Gómez-Román J, Saenz P, Molina M, Cuevas González J, Escuredo K, Santa Cruz S,
Junquera C, Simón L, Martínez A, Gutiérrez Baños J, López-Brea M, Esparza C, Val-Bernal J (2005) Fibroblast growth factor receptor 3 is overexpressed in urinary tract carcinomas and
modulates the neoplastic cell growth. _Clin Cancer Res_ 11: 459–465 PubMed Google Scholar * Halder J, Kamat A, Landen CJ, Han L, Lutgendorf S, Lin Y, Merritt W, Jennings N, Chavez-Reyes A,
Coleman R, Gershenson D, Schmandt R, Cole S, Lopez-Berestein G, Sood A (2006) Focal adhesion kinase targeting using _in vivo_ short interfering RNA delivery in neutral liposomes for ovarian
carcinoma therapy. _Clin Cancer Res_ 12: 4916–4924 Article CAS PubMed PubMed Central Google Scholar * Helander T, Carpén O, Turunen O, Kovanen P, Vaheri A, Timonen T (1996) ICAM-2
redistributed by ezrin as a target for killer cells. _Nature_ 382: 265–268 Article CAS PubMed Google Scholar * Higo M, Uzawa K, Kouzu Y, Bukawa H, Nimura Y, Seki N, Tanzawa H (2005)
Identification of candidate radioresistant genes in human squamous cell carcinoma cells through gene expression analysis using DNA microarrays. _Oncol Rep_ 14: 1293–1298 CAS PubMed Google
Scholar * Hosaka S, Nakatsura T, Tsukamoto H, Hatayama T, Baba H, Nishimura Y (2006) Synthetic small interfering RNA targeting heat shock protein 105 induces apoptosis of various cancer
cells both _in vitro_ and _in vivo_. _Cancer Sci_ 97: 623–632 Article CAS PubMed Google Scholar * Hu C, Xiong J, Zhang L, Huang B, Zhang Q, Li Q, Yang M, Wu Y, Wu Q, Shen Q, Gao Q, Zhang
K, Sun Z, Liu J, Jin Y, Tan J (2004) PEG10 activation by co-stimulation of CXCR5 and CCR7 essentially contributes to resistance to apoptosis in CD19+CD34+ B cells from patients with B cell
lineage acute and chronic lymphocytic leukemia. _Cell Mol Immunol_ 1: 280–294 CAS PubMed Google Scholar * Huang M, Mason J, Birdsey G, Amsellem V, Gerwin N, Haskard D, Ridley A, Randi A
(2005) Endothelial intercellular adhesion molecule (ICAM)-2 regulates angiogenesis. _Blood_ 106: 1636–1643 Article CAS PubMed Google Scholar * Ishigami T, Uzawa K, Higo M, Nomura H,
Saito K, Kato Y, Nakashima D, Shiiba M, Bukawa H, Yokoe H, Kawata T, Ito H, Tanzawa H (2007) Genes and molecular pathways related to radioresistance of oral squamous cell carcinoma cells.
_Int J Cancer_ 120: 2262–2270 Article CAS PubMed Google Scholar * Ivins Zito C, Kontaridis M, Fornaro M, Feng G, Bennett A (2004) SHP-2 regulates the phosphatidylinositide 3′-kinase/Akt
pathway and suppresses caspase 3-mediated apoptosis. _J Cell Physiol_ 199: 227–236 Article PubMed Google Scholar * Kowanetz M, Valcourt U, Bergström R, Heldin C, Moustakas A (2004) Id2
and Id3 define the potency of cell proliferation and differentiation responses to transforming growth factor beta and bone morphogenetic protein. _Mol Cell Biol_ 24: 4241–4254 Article CAS
PubMed PubMed Central Google Scholar * Lehmann J, Jablonski-Westrich D, Haubold U, Gutierrez-Ramos J, Springer T, Hamann A (2003) Overlapping and selective roles of endothelial
intercellular adhesion molecule-1 (ICAM-1) and ICAM-2 in lymphocyte trafficking. _J Immunol_ 171: 2588–2593 Article CAS PubMed Google Scholar * Li R, Xie J, Kantor C, Koistinen V,
Altieri D, Nortamo P, Gahmberg C (1995) A peptide derived from the intercellular adhesion molecule-2 regulates the avidity of the leukocyte integrins CD11b/CD18 and CD11c/CD18. _J Cell Biol_
129: 1143–1153 Article CAS PubMed Google Scholar * McCawley L, Matrisian L (2000) Matrix metalloproteinases: multifunctional contributors to tumor progression. _Mol Med Today_ 6:
149–156 Article CAS PubMed Google Scholar * Meineke V, Moede T, Gilbertz K, Mayerhofer A, Ring J, Köhn F, Van Beuningen D (2002) Protein kinase inhibitors modulate time-dependent effects
of UV and ionizing irradiation on ICAM-1 expression on human hepatoma cells. _Int J Radiat Biol_ 78: 577–583 Article CAS PubMed Google Scholar * Nakajima T, Yageta M, Shiotsu K, Morita
K, Suzuki M, Tomooka Y, Oda K (1998) Suppression of adenovirus E1A-induced apoptosis by mutated p53 is overcome by coexpression with Id proteins. _Proc Natl Acad Sci USA_ 95: 10590–10595
Article CAS PubMed PubMed Central Google Scholar * Okabe H, Satoh S, Furukawa Y, Kato T, Hasegawa S, Nakajima Y, Yamaoka Y, Nakamura Y (2003) Involvement of PEG10 in human
hepatocellular carcinogenesis through interaction with SIAH1. _Cancer Res_ 63: 3043–3048 CAS PubMed Google Scholar * Perez O, Kinoshita S, Hitoshi Y, Payan D, Kitamura T, Nolan G, Lorens
J (2002) Activation of the PKB/AKT pathway by ICAM-2. _Immunity_ 16: 51–65 Article CAS PubMed Google Scholar * Plowright E, Li Z, Bergsagel P, Chesi M, Barber D, Branch D, Hawley R,
Stewart A (2000) Ectopic expression of fibroblast growth factor receptor 3 promotes myeloma cell proliferation and prevents apoptosis. _Blood_ 95: 992–998 CAS PubMed Google Scholar *
Staunton D, Dustin M, Springer T (1989) Functional cloning of ICAM-2, a cell adhesion ligand for LFA-1 homologous to ICAM-1. _Nature_ 339: 61–64 Article CAS PubMed Google Scholar *
Toulany M, Kasten-Pisula U, Brammer I, Wang S, Chen J, Dittmann K, Baumann M, Dikomey E, Rodemann H (2006) Blockage of epidermal growth factor receptor-phosphatidylinositol 3-kinase-AKT
signaling increases radiosensitivity of K-RAS mutated human tumor cells _in vitro_ by affecting DNA repair. _Clin Cancer Res_ 12: 4119–4126 Article CAS PubMed Google Scholar * Tsou A,
Chuang Y, Su J, Yang C, Liao Y, Liu W, Chiu J, Chou C (2003) Overexpression of a novel imprinted gene, PEG10, in human hepatocellular carcinoma and in regenerating mouse livers. _J Biomed
Sci_ 10: 625–635 CAS PubMed Google Scholar * Zha J, Harada H, Yang E, Jockel J, Korsmeyer S (1996) Serine phosphorylation of death agonist BAD in response to survival factor results in
binding to 14-3-3 not BCL-X(L). _Cell_ 87: 619–628 Article CAS PubMed Google Scholar * Zhang H, Rao J, Guo X, Liu L, Zou T, Turner D, Wang J (2004a) Akt kinase activation blocks
apoptosis in intestinal epithelial cells by inhibiting caspase-3 after polyamine depletion. _J Biol Chem_ 279: 22539–22547 Article CAS PubMed Google Scholar * Zhang X, Chen Z, Choe M,
Lin Y, Sun S, Wieand H, Shin H, Chen A, Khuri F, Shin D (2005) Tumor growth inhibition by simultaneously blocking epidermal growth factor receptor and cyclooxygenase-2 in a xenograft model.
_Clin Cancer Res_ 11: 6261–6269 Article CAS PubMed Google Scholar * Zhang X, Ling M, Feng H, Wong Y, Tsao S, Wang X (2004b) Id-I stimulates cell proliferation through activation of EGFR
in ovarian cancer cells. _Br J Cancer_ 91: 2042–2047 Article CAS PubMed PubMed Central Google Scholar * Zhuang S, Yan Y, Daubert R, Han J, Schnellmann R (2007) ERK promotes hydrogen
peroxide-induced apoptosis through caspase-3 activation and inhibition of Akt in renal epithelial cells. _Am J Physiol Renal Physiol_ 292: F440–F447 Article CAS PubMed Google Scholar *
Zünd G, Uezono S, Stahl G, Dzus A, McGowan F, Hickey P, Colgan S (1997) Hypoxia enhances induction of endothelial ICAM-1: role for metabolic acidosis and proteasomes. _Am J Physiol_ 273:
C1571–C1580 Article PubMed Google Scholar Download references ACKNOWLEDGEMENTS We thank Lynda C Charters for editing this paper. This work was supported by a Grant-in-Aid for Japan
Society for Promotion of Science Fellows (19-52163), a Grant-in-Aid for Exploratory Research (19659520), and The 21st Century Center of Excellence (COE) Programs grant. AUTHOR INFORMATION
AUTHORS AND AFFILIATIONS * Department of Clinical Molecular Biology, Graduate School of Medicine, Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba, 260-8670, Japan T Ishigami, K Uzawa, K
Fushimi, K Saito, Y Kato, D Nakashima, M Higo, Y Kouzu & H Tanzawa * Division of Dentistry and Oral-Maxillofacial Surgery, Chiba University Hospital, 1-8-1 Inohana, Chuo-ku, Chiba,
260-8670, Japan H Bukawa & H Tanzawa * Department of Radiology, Graduate School of Medicine, Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba, 260-8670, Japan T Kawata & H Ito Authors
* T Ishigami View author publications You can also search for this author inPubMed Google Scholar * K Uzawa View author publications You can also search for this author inPubMed Google
Scholar * K Fushimi View author publications You can also search for this author inPubMed Google Scholar * K Saito View author publications You can also search for this author inPubMed
Google Scholar * Y Kato View author publications You can also search for this author inPubMed Google Scholar * D Nakashima View author publications You can also search for this author
inPubMed Google Scholar * M Higo View author publications You can also search for this author inPubMed Google Scholar * Y Kouzu View author publications You can also search for this author
inPubMed Google Scholar * H Bukawa View author publications You can also search for this author inPubMed Google Scholar * T Kawata View author publications You can also search for this
author inPubMed Google Scholar * H Ito View author publications You can also search for this author inPubMed Google Scholar * H Tanzawa View author publications You can also search for this
author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to K Uzawa. RIGHTS AND PERMISSIONS From twelve months after its original publication, this work is licensed under the
Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/ Reprints and permissions
ABOUT THIS ARTICLE CITE THIS ARTICLE Ishigami, T., Uzawa, K., Fushimi, K. _et al._ Inhibition of _ICAM2_ induces radiosensitisation in oral squamous cell carcinoma cells. _Br J Cancer_ 98,
1357–1365 (2008). https://doi.org/10.1038/sj.bjc.6604290 Download citation * Revised: 31 January 2008 * Accepted: 01 February 2008 * Published: 18 March 2008 * Issue Date: 22 April 2008 *
DOI: https://doi.org/10.1038/sj.bjc.6604290 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * oral squamous cell carcinoma * radioresistance * ICAM2
* siRNA