Play all audios:
ABSTRACT Bax and Bak are critical effectors of apoptosis. Although both are widely expressed and usually functionally redundant, recent studies suggest that Bak has particular importance in
certain cell types. Genetic and biochemical studies indicate that Bak activation is prevented primarily by Mcl-1 and Bcl-xL, whereas Bax is held in check by all pro-survival Bcl-2
homologues, including Bcl-2 itself. In this study, we have investigated whether loss of Bak or elevated Mcl-1 modulates haemopoietic abnormalities provoked by overexpression of Bcl-2. The
_Mcl-1_ transgene had little impact, probably because the expression level was insufficient to effectively reduce Bak activation. However, loss of _Bak_ enhanced lymphocytosis in
vavP-_BCL-2_ transgenic mice and increased resistance of their thymocytes to some cytotoxic agents, implying that Bak-specific signals can be triggered in certain lymphoid populations.
Nevertheless, lack of Bak had no significant impact on thymic abnormalities in vavP-_BCL-2_tg mice, which kinetic analysis suggested was due to accumulation of self-reactive thymocytes that
resist deletion. Intriguingly, although _Bak_−/− mice have elevated platelet counts, _Bak_−/−vavP-_BCL-2_ mice, like vavP-_BCL-2_ littermates, were thrombocytopaenic. To clarify why, the
vavP-_BCL-2_ platelet phenotype was scrutinised more closely. Platelet life span was found to be elevated in vavP-_BCL-2_ mice, which should have provoked thrombocytosis, as in _Bak_−/−
mice. Analysis of bone marrow chimaeric mice suggested the low platelet phenotype was due principally to extrinsic factors. Following splenectomy, blood platelets remained lower in
vavP-_BCL-2_ than wild-type mice. However, in _Rag1_−/− _BCL-2_tg mice, platelet levels were normal, implying that elevated lymphocytes are primarily responsible for _BCL-2_tg-induced
thrombocytopaenia. SIMILAR CONTENT BEING VIEWED BY OTHERS THE PHOSPHATASE PTEN LINKS PLATELETS WITH IMMUNE REGULATORY FUNCTIONS OF MOUSE T FOLLICULAR HELPER CELLS Article Open access 19 May
2022 DIFFERENTIAL IN VIVO ROLES OF MPL CYTOPLASMIC TYROSINE RESIDUES IN MURINE HEMATOPOIESIS AND MYELOPROLIFERATIVE DISEASE Article Open access 15 March 2024 MAST CELL DEFICIENCY PREVENTS
BCR::ABL1 INDUCED SPLENOMEGALY AND CYTOKINE ELEVATION IN A CML MOUSE MODEL Article Open access 09 May 2023 MAIN Apoptosis has a vital role in regulating cell numbers during haemopoiesis and
failure to remove damaged, superfluous or potentially dangerous cells can lead to malignancy or autoimmunity. Many critical life-or-death checkpoints, particularly during lymphopoiesis, are
governed by opposing factions of the Bcl-2 protein family, which regulate the ‘intrinsic’ apoptosis pathway.1, 2 Bcl-2, discovered via the t(14;18) chromosome translocation typical of human
follicular lymphoma,3, 4, 5 inhibits apoptosis,6 as do its closest homologues (Bcl-xL, Bcl-w, A1/Bfl1, Mcl-1 and, in humans, Bcl-B). Other close homologues (Bax and Bak) instead promote
apoptosis, as do distant relatives known as BH3 (Bcl-2 homology region 3)-only proteins because they share only an ∼26 amino-acid motif with the wider Bcl-2 family. During cellular stress,
BH3-only proteins are induced and bind with high avidity via their amphipathic BH3 _α_-helix to the hydrophobic groove on the surface of pro-survival Bcl-2-like proteins,7, 8, 9 thereby
preventing them from restraining any activated Bax or Bak molecules. Certain BH3-only proteins (particularly Bim and cleaved Bid) can also bind weakly and transiently to Bax and/or Bak,
triggering their conformational change and subsequent homo-oligomerisation on the outer mitochondrial membrane. As a consequence, cytochrome _c_ is released into the cytoplasm, leading to
the activation of the proteases (caspases) that provoke cellular demolition by cleaving vital proteins. Although Bax and Bak are both widely expressed and functionally redundant,10 recent
studies suggest Bak may have particular importance in certain cell types. Thus, loss of Bak results in thrombocytosis, whereas loss of Bax does not, indicating that Bak has the more
important role in regulating platelet life span.11, 12 Furthermore, loss of Bak was able to partially rescue thymic defects caused by conditional deletion of Mcl-1, whereas neither
overexpression of Bcl-2 nor loss of Bax was able to do so.13 Specificity of interactions may account for these observations: Bak binds tightly to Mcl-1 and Bcl-xL but only poorly to Bcl-2,
whereas Bax binds avidly to all the pro-survival proteins.14, 15, 16 Presumably, therefore, Bax activation can be thwarted by all pro-survival proteins, including Bcl-2, whereas Bak is kept
in check by Mcl-1 and Bcl-xL. To further explore the role of Bak-specific cell death during haemopoiesis, we have investigated whether loss of Bak or increased expression of Mcl-1 enhances
the impact of pan-haemopoietic overexpression of Bcl-2. To do so, we crossed vavP-_BCL-2_ transgenic (hereafter _BCL-_2tg) mice17 with _Bak_−/−10 or vavP-_Mcl-1_ transgenic (hereafter
_Mcl-1_tg) mice18 and compared the phenotypes of single and doubly mutant offspring. This study also enabled us to undertake further analysis of the puzzling thymic and platelet phenotype of
_BCL-2_tg mice. RESULTS IMPACT OF OVEREXPRESSION OF MCL-1 AND LOSS OF BAK ON HAEMOPOIESIS IN _BCL-2TG_ MICE Overexpression of Bcl-2 via the haemopoietic cell-specific vavP-driven
transgene19 enhances the survival of T- and B-lymphoid cells, which accumulate in excessive numbers in the periphery.17, 20 In addition, _BCL-2_tg mice have a distinctive thymic phenotype: a
reduced proportion of pre-T cells (CD4+CD8+ double positive; hereafter DP) and elevated proportions of the other three major populations (CD4−CD8− double negative, hereafter DN; CD4+CD8−
single positive, hereafter CD4SP; and CD4−CD8+ single positive, hereafter CD8SP).17 This is referred to as the ‘low DP’ thymic phenotype of _BCL-2_tg mice (see further below). To assess
whether overexpression of Mcl-1 exacerbates the _BCL-2_tg phenotype, we compared the composition of blood and haemopoietic tissues in 6-week-old neonates. Despite a minor (_P_≤0.05) increase
in the total number of DP thymocytes in _Mcl-1/BCL-2_ bi-transgenic compared with _BCL-2_tg mice, the _proportion_ of DP thymocytes was comparably low (lower left panel in Supplementary
Figure 1B) and there was little impact on the _BCL-2_tg-induced lymphocytosis in the other lymphoid organs (Supplementary Figure 1 and Supplementary Table 1). To assess the impact of loss of
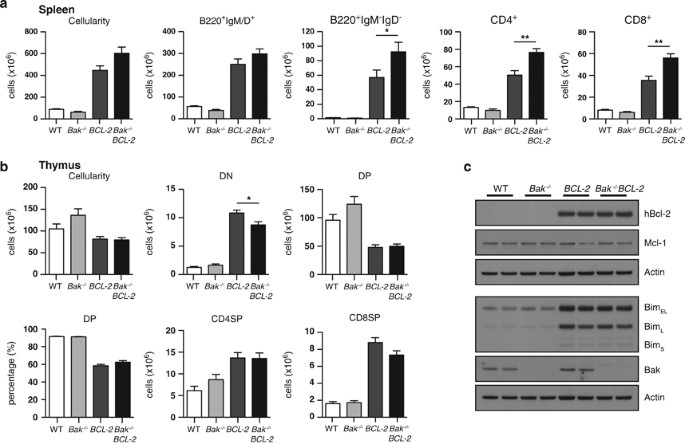
Bak, we first analysed haemopoietic tissues in young adult (12- to 14-week-old) mice. Loss of Bak had little impact alone, but did increase lymphocytosis in _BCL-2_tg mice (Figure 1 and
Supplementary Table 2). This was most apparent in the spleen where cellularity, already elevated approximately fivefold in _BCL-2_tg mice, increased to nearly sevenfold in _Bak_−/− _BCL-2_tg
animals, due primarily to a further elevation in the number of immunoglobulin (Ig) isotype-switched (B220+IgM−IgD−) B-lymphoid cells and mature T cells (CD4+ and CD8+; Figure 1a). A
comparable cross of _Bax_−/− and _BCL-2_tg mice revealed no comparable differences between _BCL-2_tg and _Bax_−/− _BCL-2_tg animals (Supplementary Table 3). Lack of Bak did not notably alter
the low proportion of DP thymocytes in the _BCL-2_tg thymus (Figure 1b and Supplementary Table 2). _BCL-2_tg thymocytes have very high levels of the pro-apoptotic BH3-only protein Bim, most
being sequestered by BCL-2,21 and Bim levels were unchanged in either _Bak_−/− _BCL-2_tg or _Mcl-1_tg_/BCL-2_tg mice (Figure 1c and Supplementary Figure 1C). To explore the perturbation of
T lymphopoiesis more closely, we compared the thymus and spleen of _BCL-2_tg and _Bak_−/− _BCL-2_tg mice at three time points: 6–8, 12 and 24 weeks (Figure 2 and Supplementary Table 4). The
decrease in the number of DP thymocytes was not as marked at 6–8 weeks (∼80 % that in wild-type (WT) littermates) as at 12 and 24 weeks (∼46% and ∼47%, respectively), although mature
thymocytes (CD4SP and CD8SP) were already significantly elevated at 6–8 weeks, as were T cells in the spleen. Strikingly, there was an approximately fourfold increase in the DN population in
6- to 8-week-old _BCL-2_tg and _Bak_−/−_BCL-2_tg mice, and this was almost entirely due to T-cell receptor _β_+ (TCR_β_+) DN cells (elevated 16-fold; Figure 2a). These cells are probably
mature T cells (TCR_β_ is highly expressed) that have downregulated their co-receptors (CD4 or CD8) because they are autoreactive and have escaped negative selection.22 The early increase in
these ‘pseudo DN’ cells may inhibit the production of _bona fide_ DN thymic progenitor cells (see Discussion). LOSS OF BAK INCREASES RESISTANCE OF _BCL-2_TG THYMOCYTES TO CERTAIN APOPTOTIC
STIMULI To compare the resistance of thymocytes of the different genotypes to apoptosis, we first performed _in vitro_ tests. In the absence of cytokines, the _Mcl-1_ and _BCL-2_ transgenes
provided comparable protection, but in the presence of cytotoxic agents, the _BCL-2_ transgene provided greater protection, consistent with previous observations,23 and co-expression of both
conferred no additional advantage (Supplementary Figure 2). By itself, loss of Bak did not protect DP thymocytes from spontaneous death in culture or from apoptosis induced by dexamethasone
or phorbol 12-myristate-13-acetate (PMA), but it did enhance resistance to ionomycin at low (1 _μ_g/ml) concentration and, at early time points, to DNA damage (_γ_-irradiation and
etoposide; Figure 3a and Supplementary Figure 3). Furthermore, loss of Bak enhanced the resistance of DP thymocytes expressing the _BCL-2_ transgene to apoptosis induced by etoposide
treatment and, modestly, to _γ_-irradiation (Figure 3a). DP thymocytes are exquisitely sensitive to CD3 antibody, which triggers apoptosis by aggregating the TCR-CD3 complex.24, 25 As a
further test, therefore, we injected mice of all four genotypes with an optimal dose of CD3_ɛ_ monoclonal antibody (mAb) or with an Ig isotype-matched control antibody, and determined thymus
weight and cellularity after 40 h (Figure 3b). Following CD3 antibody treatment, thymus weight dropped substantially in both WT and _Bak_−/− mice, due primarily to a reduction in DP
thymocytes. As reported previously,26 expression of the _BCL-2_ transgene provided DP thymocytes with only partial protection against this insult (65%). Of note, however, protection in the
_Bak_−/− _BCL-2_tg mice was considerably more robust (>90%), suggesting that TCR-activated apoptosis involves a Bak-specific component. PATHOLOGY _BCL-2_tg mice are predisposed to the
development of autoimmune kidney disease and, in later life, follicular lymphoma.20 To determine whether overexpression of Mcl-1 or loss of Bak increased the risk of morbidity, cohorts of
mice were monitored for 12 months. Most mice that became ill during this period showed evidence of autoimmune kidney disease: weight loss, blood in the urine and pale, often speckled,
kidneys. The overall survival of _Mcl-1_tg_/BCL-2_tg mice over 12 months was comparable to that of _BCL-2_tg mice and serum Ig levels were similarly elevated in both cohorts (Supplementary
Figure 4). The autoimmune kidney disease was most likely due to overexpression of Bcl-2 as _Mcl-1_tg mice do not develop comparable symptoms.18 Loss of Bak resulted in a trend towards
earlier morbidity in _BCL-2_tg mice, although the difference between _Bak_−/−_BCL-2_tg and _BCL_-2tg animals did not achieve statistical significance (Supplementary Figure 5A). The elevation
in IgM, IgG and IgA antibody-secreting cells (ASCs) in the spleen and bone marrow was similar between these cohorts (Supplementary Figures 5B and C). Although a few lymphomas developed in
transgenic mice of each genotype, neither overexpression of Mcl-1 nor loss of Bak enhanced the susceptibility of the _BCL-2_tg mice to follicular lymphoma during the time period analysed.
Like _Mcl-1_tg mice,18 certain _Mcl-1_tg_/BCL-2_tg mice developed other lymphomas, at comparable frequency. IMPACT ON PLATELETS Both _Bak_−/− and _BCL-2_tg mice have platelet abnormalities:
loss of Bak causes thrombocytosis,11, 12 but Bcl-2 overexpression results, surprisingly, in thrombocytopaenia.17 We therefore determined platelet levels in the blood of _Bak_−/−_BCL-2_tg
mice. Unexpectedly, loss of Bak did not elevate platelets in the blood of _BCL-2_tg mice; rather, _Bak_−/− _BCL-2_tg mice, like _BCL-2_tg mice, were thrombocytopaenic (Figure 4a). Thus,
Bcl-2 overexpression is dominant over loss of Bak in regard to this phenotype. At steady state, platelets circulate in the blood for up to 5 days in mice, 10 days in humans27, 28 and the
elevated platelet count in _Bak_−/− mice is due to increased platelet life span.11 To determine the impact of overexpression of BCL-2 on platelet life span, we injected _BCL-2_tg,
_Bak_−/−_BCL-2_tg and _Bak_−/− mice with NHS-biotin and tracked the disappearance of labelled platelets from the circulation (Figure 4b). Consistent with previous reports,11 platelet life
span was significantly extended in _Bak_−/− mice, with a half-life of 100 h compared with 60 h in WT littermates. Importantly, _BCL-2_tg animals showed an intermediate platelet half-life of
75 h. Thus, overexpression of Bcl-2 enhances the life span of platelets, albeit more modestly than loss of Bak. Underlining the fundamental importance of Bak in regulating platelet life
span, there was no additive effect in _Bak_−/−_BCL-2_tg mice; platelet survival in these animals was indistinguishable from that seen in the _Bak_−/− cohort. Megakaryocyte (not shown) and
platelet counts were normal in young _Mcl-1_tg mice18 (Supplementary Figures 6A and B) and platelet life span was also normal (Supplementary Figure 6C), suggesting that the modest increase
in platelet Mcl-1 levels in this model (Supplementary Figure 6D), coupled with its short half-life,29 is not sufficient to have any measurable effect on overall survival times. Like
_BCL-2_tg mice, _Mcl-1_tg/_BCL-2_tg mice were thrombocytopaenic, although the deficit in mature platelets was not quite as profound (Supplementary Figures 6A and B). WHY DOES HAEMOPOIETIC
OVEREXPRESSION OF BCL-2 CAUSE THROMBOCYTOPAENIA? The thrombocytopaenia in _BCL-2_tg and _Bak_−/−_BCL-2_tg mice (Figure 4a) is at odds with the enhanced platelet life span in these animals
(Figure 4b). To clarify this conundrum, we decided to investigate the _BCL-2_tg platelet phenotype in more detail. Megakaryocyte frequency was normal in the bone marrow of _BCL-2_tg mice, as
in _Bak_−/− mice, albeit somewhat increased in _Bak_−/−_BCL-2_tg mice (Figure 4c). Although the percentage of reticulated (newly formed) platelets was unchanged in _BCL-2_tg compared with
WT mice, their absolute number was significantly reduced (Figure 4d), implying a defect in platelet production from megakaryocytes. We therefore tested the ability of megakaryocytes from WT
and _BCL-2_tg mice to produce pro-platelets _in vitro_. Consistent with a previous report,30 Bcl-2 overexpression did not decrease proplatelet formation by fetal liver-derived megakaryocytes
but, rather, caused a small but significant increase (Figure 4e). These data indicate that, at least in culture, _BCL-2_tg megakaryocytes exhibit no obvious defects in maturation and
proplatelet formation. Nevertheless, when we analysed the ability of mice to recover from acute thrombocytopaenia induced by injection of anti-platelet serum (APS), we found that platelet
rebound was significantly impaired in _BCL-2 versus_ WT mice (Figure 4f). To further clarify the basis for the platelet defect, we examined whether thrombocytopaenia in _BCL-2_tg mice is
platelet-intrinsic or -extrinsic (Figure 5). Bone marrow chimaeric mice were generated by injecting lethally irradiated Ly5.1 mice with 2 × 106 Ly5.2 bone marrow cells from _GFP_tg mice31
(in which GFP expression is ubiquitous) or _BCL-2_tg mice, or with a 50 : 50 mixture of both. Analysis at 9 weeks post-transplantation showed that the red blood cell count was comparable in
all three classes of reconstituted mice but, as expected, the white blood cell count was higher and the platelet count was lower in mice reconstituted with _BCL-2_tg cells than in those
reconstituted solely with _GFP_tg cells (Figures 5a–c). In the _GFP_tg/_BCL-2_tg chimaeras, where the proportion of GFP+ platelets was around 40% (Figure 5d and Supplementary Table 5), total
platelet counts were equivalent to those seen in mice reconstituted solely with _BCL-2_tg cells (Figure 5c). By 12 weeks, there were 24% GFP-positive platelets and the proportion of
GFP-positive megakaryocytes was similar (22%; Supplementary Table 5). We infer that (i) the _BCL-2_tg-driven thrombocytopaenia derives from bone marrow cells; (ii) WT (_GFP_tg) haemopoietic
cells cannot compensate for the defect; and (iii) the defect in platelets is cell extrinsic. Ten weeks post-reconstitution, mice were injected with APS and platelet numbers assessed daily
for 5 days (Figure 5e). In all mice, APS reduced circulating platelet numbers to <10% within 24 h. In the chimaeric _GFP_tg/_BCL-2_tg mice, the rebound was more robust than that seen in
_BCL-2_tg-reconstituted mice, but still significantly impaired relative to _GFP_tg-reconstituted counterparts (Figure 5e). If the rate of production of platelets had been slower only in
cells expressing the _BCL-_2 transgene, then the proportion of GFP-positive (i.e., _BCL-2_tg negative) platelets would have been expected to increase over time in the chimaeric
_GFP_tg/_BCL-2_tg mice. However, it remained constant over the 5 days (Figure 5f), again suggesting that megakaryocyte- and platelet-extrinsic factors underlie the thrombocytopaenia observed
in _BCL-2_tg animals. ROLE OF THE SPLEEN As the spleen can increase or decrease the pool of platelets that it sequesters,32 and _BCL-2_tg (and _Bak_−/−_BCL-2_tg) mice have enlarged spleens
(Supplementary Table 2), we determined platelet counts before, and 4 weeks post, splenectomy in both WT and _BCL-2_tg mice. Although platelet numbers increased in both cohorts, the relative
increment was equivalent: 1.4-fold increase in WT and 1.5-fold in _BCL-2_tg mice (Figure 6a). Furthermore, when acute thrombocytopaenia was induced in splenectomised mice, _BCL-2_tg animals
mounted a significantly impaired response, with platelet counts very significantly lower than those seen in WT littermates 5 days post induction (Figure 6b). Although the degree of
thrombocytopaenia in the splenectomised _BCL-2_tg animals is modest, when the increase in platelet life span in these animals is taken into account, it indicates a significant residual
defect, which is amplified under conditions of emergency thrombopoiesis (Figure 6b). ROLE OF LYMPHOCYTES Having established that the thrombocytopaenia in _BCL-2_tg mice was due to
environmental factors involving bone marrow-derived cells but extrinsic to megakaryocytes and platelets, we reasoned that a non-myeloid cell might be playing a role. We therefore crossed the
_BCL-2_tg mice with _Rag1_−/− mice, which lack mature B and T lymphocytes.33 Circulating lymphocytes were greatly reduced in _BCL-2_tg _Rag1_−/− compared with _BCL-2_tg mice (Figure 6c), as
expected, but blood platelet counts were elevated 1.7-fold (Figure 6d). Indeed, platelet counts in _BCL-2_tg _Rag1_−/− mice were comparable to those in WT and _Rag1_−/− mice. Thus, the
absence of mature lymphocytes prevents thrombocytopaenia in _BCL-2_tg mice. Given the increased platelet life span conferred by overexpression of BCL-2, however, the increase is not as much
as might be expected, suggesting that additional (unknown) factors might contribute to the _BCL-2_tg phenotype. DISCUSSION Bax and Bak are the critical effectors of apoptosis. Although both
are widely expressed and functionally redundant, genetic and biochemical data suggest that Bak is held in check primarily by Mcl-1 and Bcl-xL, whereas Bax activation is inhibited by all five
pro-survival Bcl-2 homologues.14, 15, 16 Recent studies have suggested that Bak activation is the principal trigger for apoptosis in platelets11 and in certain thymic populations.13 To test
whether Bak is critical at other stages of haemopoietic development, we crossed vavP-_BCL-2_tg mice with mice lacking Bak. We also crossed vavP-_BCL-2_tg mice with vavP-_Mcl-1_tg mice, in
which Bak activation in haemopoietic cells should be more difficult to achieve than in WT mice. Constitutive overexpression of Bcl-2 during haemopoiesis results in many changes: thymic
abnormalities, most notably a low proportion of DP thymocytes; striking peripheral lymphocytosis involving both B- and T-lymphoid cells; elevated ASCs and serum Ig; poorly understood
thrombocytopaenia; and predisposition to autoimmune kidney disease and late onset follicular lymphoma.19, 20 Our analysis of young _Mcl-1_tg/_BCL-2_tg mice suggested that overexpression of
Mcl-1 did not significantly modulate the _BCL-2tg_ phenotype (Supplementary Figure 1), probably because the level of Mcl-1, albeit significantly higher than normal,18 was insufficient to
effectively reduce Bak activation. In contrast, _Bak_−/−_BCL-2_tg mice developed a more severe lymphocytosis than _BCL-2_tg littermates, primarily due to a further elevation in the numbers
of mature B- and T-lymphoid cells (Figure 1a), changes that were not seen in _Bax_−/−_BCL-2_tg mice (Supplementary Table 3). The implication is that Bak can have a non-redundant role in
maintaining homeostasis within these populations. Bak must be less critical in regulating ASCs, however, because their frequency was comparable in _BCL-2_tg and _Bak_−/−_BCL-2_tg mice, as
was the risk of autoimmune kidney disease (Supplementary Figure 5). Lack of Bak had no impact on the low proportion of DP thymocytes characteristic of _BCL-2_tg mice (Figure 1b), which
kinetic analysis suggested was preceded by an accumulation of DN cells expressing high levels of TCR_β_ (Figure 2a). Also notable in _Bim_−/− and _Bim_−/−_Puma_−/− mice, these TCR_β_+DN
cells are thought to be thymocytes that have downregulated their co-receptors26, 34 after escaping negative selection at the DP stage.22 The ‘pseudo’ DN cells may inhibit the expansion and
maturation of _bona fide_ DN progenitor cells to DP (pre-T) cells,22 perhaps due to competition for niche cells or cytokines. DP thymocytes, normally highly sensitive to apoptosis, are
rendered significantly resistant to most cytotoxic agents by overexpression of Bcl-2.24 Loss of Bak modestly enhanced the resistance of DP thymocytes from _BCL-2_tg mice to apoptosis induced
by DNA-damaging agents _in vitro_ (Figure 3a) and significantly increased their resistance to CD3 antibody treatment _in vivo_ (Figure 3b). Thus, Bak may be non-redundant for effecting
apoptosis of certain lymphoid populations, perhaps because of kinetic differences in the activation of Bak _versus_ Bax and/or a Bak-specific component of certain apoptosis-inducing signals.
Ca+ flux (which is induced by TCR/CD3 ligation) may have an important role in the Bak activation, as, by itself, loss of Bak provided significant protection against apoptosis induced by
ionomycin (Figure 3a). We have shown previously that loss of the pro-apoptotic BH3-only protein Bim also provides greater resistance to CD3 antibody than the _BCL-2_ transgene and that Bim
is the critical downstream apoptosis effector.26, 35 Bim can activate both Bak and Bax as well as inhibit all pro-survival Bcl-2 family members.36, 37, 38 Our most unexpected finding
involved platelets. The observation that _BCL-2_tg mice are thrombocytopaenic17 led to conjecture that platelet shedding by megakaryocytes involves activation of apoptosis,39 and a
considerable body of subsequent work supported that notion (reviewed in White and Kile32). However, counter to this proposal, we and others recently demonstrated that deletion of Bak and Bax
does not impair platelet production.12, 40, 41 Thus, an explanation for the thrombocytopaenic phenotype of _BCL-2_tg mice has remained elusive. Platelet survival is primarily dependent on
Bcl-xL, which is required in ageing platelets to restrain Bak, the principal mediator of their apoptosis.11, 12, 42 We found that platelet life span is extended in _BCL-2_tg mice (Figure
4b), demonstrating for the first time that overexpression of a Bcl-2 family pro-survival protein can positively influence the survival of platelets in the circulation. BCL-2 may be able to
partly restrain Bak at these high, non-physiological concentrations, as well as inhibit normal targets Bax and Bad, which also influence platelet life span,12, 43 albeit not to nearly the
same extent as loss of Bak.12 In any event, the thrombocytopaenia in _BCL-2_tg animals must actually be more severe than suggested by the platelet count alone – were this not the case, the
extended platelet life span conferred by the _BCL-2_ transgene would have provoked mild thrombocytosis. Consistent with Kozuma and colleagues,30 megakaryocyte numbers were normal in
_BCL-2_tg mice, and their ability to form pro-platelets _in vitro_ was unimpaired. However, contrary to that study, removal of the spleen did not raise their platelet counts to that found in
splenectomised WT mice, either at steady state or during emergency thrombopoiesis (Figures 6a and b), ruling out increased splenic sequestration as the principal cause. Instead, we believe
the thrombocytopaenia is underpinned by a defect in platelet shedding caused by extrinsic factors in the _BCL-2_tg haemopoietic microenvironment. Analysis of bone marrow chimaeras supported
this notion, as WT megakaryocytes in a _BCL-2_tg environment were unable to produce platelets at the same rate as those in a WT environment (Figure 5e). Our data suggest that the
lymphocytosis induced by the _BCL-2_ transgene is a major contributing factor, as platelet counts increased to WT levels in a _Rag1_−/− background (Figure 6d). Other as yet unidentified
haemopoietic cells must also have a role, however, as platelet counts were not elevated above WT levels, as would be expected from the increased platelet life span conferred by
overexpression of BCL-2. As vavP-_BCL-2_ transgene expression also elevates levels of bone marrow macrophages and monocytes (Supplementary Table 2), these cells may contribute. However,
whether the impairment of platelet production is mediated directly by lymphoid and other haemopoietic cell types or indirectly, via effects on other tissues, remains to be determined.
Furthermore, it remains formally possible that, in addition, BCL-2 overexpression does have a subtle intrinsic impact on megakaryocyte function. Specific overexpression of BCL-2 in the
megakaryocyte lineage _in vivo_ would enable this to be assessed in the absence of extrinsic factors. MATERIALS AND METHODS MICE All mice used were on a C57BL/6J background and bred at the
Walter and Eliza Hall Institute (WEHI). Experimental protocols were approved by Animal Ethics Committee of WEHI. Transgenic mouse lines were vavP-_Mcl-1_(33)18 and vavP-_BCL-2_(69),17, 20
which, respectively, express FLAG-tagged mouse Mcl-1 protein and human BCL-2 protein in haemopoietic cells.19 To generate _Mcl-1_tg/_BCL-2_tg mice, _Mcl-1_tg males were mated with _BCL-2_tg
females. To generate _Bak_−/−_BCL-2_tg mice, _BCL-2_tg males were crossed with _Bak_−/−10 females, then _Bak_+/− _BCL-2_tg males were mated with _Bak_+/− females. Crosses were also performed
with _Rag1_−/−,44 _Bax_+/−45 and Tg(UBC-GFP)30Scha/J mice (_GFP_tg mice).31 HAEMOPOIETIC ANALYSIS Single-cell suspensions were prepared from spleen, lymph nodes, bone marrow and thymus and
viable leukocytes enumerated using a haemocytometer and trypan blue exclusion or with a CASY Cell Counter (Scharfe System GmbH, Reutlingen, Germany). An ADVIA 2120 haematology analyser
(Siemens, Erlangen, Germany) was used for obtaining blood cell counts. The remaining blood was depleted of red cells by treatment with 0.168 M ammonium chloride before fluorescence-activated
cell sorting (FACS) analysis. Cell composition was determined by staining with fluorochrome-labelled surface marker-specific monoclonal antibodies followed by FACS analysis using an LSRI
(BD Biosciences, Franklin Lakes, NJ, USA). Data were processed using FlowJo Version 9.3.2 (TreeStar, Ashland, OR, USA) and Weasel Version 3.0 software (Walter and Eliza Hall Institute,
Melbourne, VIC, Australia). The monoclonal antibodies, produced and labelled with fluorescein isothiocyanate, R-phycoerythrin or allophycocyanin (APC) at WEHI unless otherwise indicated,
were: RB6-8C5, anti-Gr1; MI/70, anti-Mac1; H129.19, anti-CD4; YTS169, anti-CD8; Ter119, anti-erythroid marker; ID3, anti-CD19; RA3-6B2, anti-CD45R-B220; 5.1, anti-IgM; 11-26C, anti-IgD; S7,
anti-CD43; T24-31, anti-Thy1; H57-59, anti-TCR_β_; Jo2, anti-Fas/CD95 (BD Biosciences); anti-PNA (Vector Laboratories, Cambridgeshire, UK). Blood platelet counts were determined using an
ADVIA 2120 analyser (Siemens) or by flow cytometry, as follows: 5 _μ_l of tail vein blood was diluted 40-fold in PBS in an EDTA tube and stained with CD41 antibody (clone MWReg30, BD
Biosciences) labelled with APC; 10 _μ_l was then added to PBS (980 _μ_l) containing 10 _μ_l beads (ProSciTech, Townsville, QLD, Australia; 3.5–4 _μ_m beads, 1 × 107/ml) and 1000 events were
acquired by flow cytometry; platelet frequency was calculated as dilution factor (4000) × number of events in platelet gate (CD41+) × 1 × 105. Reticulated platelet numbers were determined by
staining with thiazole orange and CD41-APC antibody11 followed by analysis on a FACSCalibur flow cytometer (BD Biosciences). Platelet life span was investigated by _in vivo_ labelling with
biotin.11 Induction of thrombocytopaenia with APS, culture of fetal liver megakaryocytes, proplatelet formation assays and platelet preparation were performed as described.11, 12 CD3
ANTIBODY TREATMENT Mice were injected intraperitoneally with 30 _μ_g hamster mAb to mouse CD3_ɛ_ (145-2C11) or, as a control, isotype-matched antibody to TCR_γ_ (GL3) and killed after 40 h
for analysis of thymic cellularity and composition by flow cytometry. SURVIVAL ASSAYS Thymocyte populations isolated by flow cytometry were cultured at 0.2–0.5 × 106 cells/ml in high-glucose
Dulbecco’s Modified Eagle’s medium supplemented with 10% fetal calf serum (Bovogen, Melbourne, VIC, Australia), 50 _μ_M 2-mercaptoethanol (Sigma-Aldrich, St. Louis, MO, USA) and 100 _μ_M
asparagine (Sigma-Aldrich) without additional cytokines in the presence or absence of 10 _μ_g/ml etoposide (Pfizer, Sydney, NSW, Australia), 10 _μ_M dexamethasone phosphate (Hospira, Lake
Forest, IL, USA), 10 ng/ml PMA (Sigma-Aldrich), 10 _μ_g/ml ionomycin (Sigma-Aldrich) or following treatment with 10 Gy _γ_-irradiation. Cell viability was determined by flow cytometry after
staining with fluorescein isothiocyanate-conjugated Annexin V and propidium iodide. Specific viability was calculated at each time point as (viability of treated cells/viability of untreated
cells) × 100%. Alternatively, apoptotic cells were identified by active caspase-3 staining. Cells were fixed and permeabilised using the BD Cytofix/Cytoperm Kit for 20 min, then washed with
BD Perm/Wash buffer and stained with phycoerythrin rabbit anti-active caspase-3 antibody (clone C92-605, BD Biosciences), then washed again in BD Perm/Wash buffer before analysing by flow
cytometry. HAEMOPOIETIC RECONSTITUTIONS The _GFP_tg/_BCL-2_tg haemopoietic chimaeras were generated using bone marrow collected from _GFP_tg and _BCL-2_tg mice. Bone marrow (2 × 106) cells
suspended in phosphate-buffered saline were injected into lethally irradiated (2 × 5.5 Gy spaced by 2 h) C57BL/6-CD45.1 (Ly5.1) mice. To prevent infections, transplanted animals were
initially provided with water containing neomycin (Sigma-Aldrich). After stable reconstitution of their haemopoietic system (9 weeks later), blood was collected from the retro-orbital plexus
for ADVIA and FACS analysis (see above). WESTERN BLOT ANALYSIS Thymocyte lysates were prepared using RIPA buffer (300 mM NaCl, 2% octylphenoxypolyethoxyethanol (IGEPAL CA-630;
Sigma-Aldrich), 1% deoxycholic acid, 0.2% SDS, 100 mM Tris-HCl pH 8.0) containing protease inhibitors (Roche, Basel, Switzerland) and platelet lysates with NP40 lysis buffer (1%
octylphenoxypolyethoxyethanol, 150 mM NaCl, 50 mM Tris-HCl, pH 7.4) containing protease inhibitors. Proteins were separated on NuPAGE Bis-Tris gels (Life Technologies, Carlsbad, CA, USA)
according to manufacturer’s instructions. Blots were probed with: anti-Mcl-1 (clone 19C4-15; WEHI mAb facility), anti-human Bcl-2 (clone Bcl-2-100;46 WEHI mAb facility), anti-Bcl-2 (clone 7;
BD Biosciences), anti-Bim (polyclonal; Enzo Lifesciences, Farmingdale, NY, USA), anti-Bcl-xL (polyclonal; BD Biosciences), anti-Bak (polyclonal; Sigma-Aldrich) and anti-_β_-actin (clone
AC-74; Sigma-Aldrich). ELISA AND ENZYME-LINKED IMMUNOSPOT (ELISPOT) ASCs were enumerated by ELISPOT. MultiScreen-HA filter plates (Merck Millipore, Billerica, MA, USA ) were coated with 2
_μ_g/ml anti-mouse total Ig (DA, Silenus Laboratories, Boronia, VIC, Australia) or 10 _μ_g/ml anti-mouse IgA (Southern Biotech, Birmingham, AL, USA). Red cell-depleted spleen or bone marrow
cells were added at 1 × 104 or 1 × 105 per well in RPMI/5% FCS/5 _μ_M 2-mercaptoethanol and incubated for 18–19 h. The plates were washed and incubated with secondary antibodies: anti-mouse
IgA-biotin, anti-mouse IgG1-HRP/IgG2a-HRP/IgG2b-biotin/IgG3-HRP or anti-mouse IgM-HRP (Southern Biotech). For biotinylated antibodies, plates were washed again and incubated with
streptavidin-HRP (Southern Biotech). ELISPOTs were revealed by the addition of substrate solution: 250 _μ_g/ml 3-amino-9-ethylcarbazole (Sigma-Aldrich) in 0.05 M sodium acetate (pH 5.0) and
0.03% H2O2. ELISPOTs were counted on an ELISPOT reader (Autoimmun Diagnostika GMBH, Strasburg, Germany). Serum Ig levels were determined by ELISA. Plates were coated with specific anti-mouse
Ig antibodies (Southern Biotech). Purified mouse monoclonal IgM_κ_ (TEPC 183), IgG1_κ_ (MOPC31c), IgG2a_κ_ (UPC 10), IgG2b_κ_ (MOPC 141), IgG3_λ_ (Y5606) and IgA_λ_ (MOPC315; Sigma-Aldrich)
were used to quantify Ig concentration. Biotinylated or HRP-conjugated isotype-specific antibodies were as for ELISPOT assay (with streptavidin-HRP, if required). The assay was developed
with 0.54 mg/ml diammonium 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) (Sigma-Aldrich) in 0.1 M citric acid (pH 4.4) and 0.03% H2O2. Absorbance at 492 nm was measured in a
microplate reader. STATISTICAL ANALYSIS GraphPad Prism (Version 5.0a; GraphPad Software, La Jolla, CA, USA) was used to graph and statistically analyse data. The Mann–Whitney test was used
to determine statistical significance when samples had a clear difference in S.D., otherwise, an unpaired two-tailed _t_-test was performed. For analysis of Kaplan–Meier mouse survival
curves, significance was determined using the log-rank (Mantel–Cox) test. ABBREVIATIONS * APS: anti-platelet serum * ASC: antibody-secreting cell * BH: Bcl-2 homology * DN: double negative *
DP: double positive * FACS: fluorescence-activated cell sorting * Ig: immunoglobulin * mAb: monoclonal antibody * PMA: phorbol 12-myristate-13-acetate * SP: single positive * TCR: T-cell
receptor * tg: transgenic * WT: wild type REFERENCES * Strasser A, Cory S, Adams JM . Deciphering the rules of programmed cell death to improve therapy of cancer and other diseases. _EMBO J_
2011; 30: 3667–3683. Article CAS Google Scholar * Llambi F, Moldoveanu T, Tait SW, Bouchier-Hayes L, Temirov J, McCormick LL _et al_. A Unified Model of Mammalian BCL-2 Protein Family
Interactions at the Mitochondria. _Mol Cell_ 2011; 44: 517–531. Article CAS Google Scholar * Tsujimoto Y, Finger LR, Yunis J, Nowell PC, Croce CM . Cloning of the chromosome breakpoint of
neoplastic B cells with the t(14;18) chromosome translocation. _Science_ 1984; 226: 1097–1099. Article CAS Google Scholar * Cleary ML, Sklar J . Nucleotide sequence of a t(14;18)
chromosomal breakpoint in follicular lymphoma and demonstration of a breakpoint-cluster region near a transcriptionally active locus on chromosome 18. _Proc Natl Acad Sci USA_ 1985; 82:
7439–7443. Article CAS Google Scholar * Bakhshi A, Jensen JP, Goldman P, Wright JJ, McBride OW, Epstein AL _et al_. Cloning the chromosomal breakpoint of t(14;18) human lymphomas:
clustering around JH on chromosome 14 and near a transcriptional unit on 18. _Cell_ 1985; 41: 899–906. Article CAS Google Scholar * Vaux DL, Cory S, Adams JM . _Bcl_-2 gene promotes
haemopoietic cell survival and cooperates with c-_myc_ to immortalize pre-B cells. _Nature_ 1988; 335: 440–442. Article CAS Google Scholar * Sattler M, Liang H, Nettesheim D, Meadows RP,
Harlan JE, Eberstadt M _et al_. Structure of Bcl-xL-Bak peptide complex: recognition between regulators of apoptosis. _Science_ 1997; 275: 983–986. Article CAS Google Scholar * Petros AM,
Nettseheim DG, Wang Y, Olejniczak ET, Meadows RP, Mack J _et al_. Rationale for Bcl-xL/Bad peptide complex formation from structure, mutagenesis, and biophysical studies. _Protein Sci_
2000; 9: 2528–2534. Article CAS Google Scholar * Liu X, Dai S, Zhu Y, Marrack P, Kappler JW . The structure of a Bcl-xL/Bim fragment complex: Implications for Bim function. _Immunity_
2003; 19: 341–352. Article CAS Google Scholar * Lindsten T, Ross AJ, King A, Zong W, Rathmell JC, Shiels HA _et al_. The combined functions of proapoptotic Bcl-2 family members Bak and
Bax are essential for normal development of multiple tissues. _Mol Cell_ 2000; 6: 1389–1399. Article CAS Google Scholar * Mason KD, Carpinelli MR, Fletcher JI, Collinge JE, Hilton AA,
Ellis S _et al_. Programmed anuclear cell death delimits platelet life span. _Cell_ 2007; 128: 1173–1186. Article CAS Google Scholar * Josefsson EC, James C, Henley KJ, Debrincat MA,
Rogers KL, Dowling MR _et al_. Megakaryocytes possess a functional intrinsic apoptosis pathway that must be restrained to survive and produce platelets. _J Exp Med_ 2011; 208: 2017–2031.
Article CAS Google Scholar * Dunkle A, Dzhagalov I, He YW . Mcl-1 promotes survival of thymocytes by inhibition of Bak in a pathway separate from Bcl-2. _Cell Death Differ_ 2010; 17:
994–1002. Article CAS Google Scholar * Willis SN, Chen L, Dewson G, Wei A, Naik E, Fletcher JI _et al_. Proapoptotic Bak is sequestered by Mcl-1 and Bcl-xL, but not Bcl-2, until displaced
by BH3-only proteins. _Gene Dev_ 2005; 19: 1294–1305. Article CAS Google Scholar * Willis SN, Fletcher JI, Kaufmann T, van Delft MF, Chen L, Czabotar PE _et al_. Apoptosis initiated when
BH3 ligands engage multiple Bcl-2 homologs, not Bax or Bak. _Science_ 2007; 315: 856–859. Article CAS Google Scholar * Uren RT, Dewson G, Chen L, Coyne SC, Huang DCS, Adams JM _et al_.
Mitochondrial permeabilization relies on BH3 ligands engaging multiple pro-survival Bcl-2 relatives, not Bak. _J Cell Biol_ 2007; 177: 277–287. Article CAS Google Scholar * Ogilvy S,
Metcalf D, Print CG, Bath ML, Harris AW, Adams JM . Constitutive bcl-2 expression throughout the hematopoietic compartment affects multiple lineages and enhances progenitor cell survival.
_Proc Natl Acad Sci USA_ 1999; 96: 14943–14948. Article CAS Google Scholar * Campbell KJ, Bath ML, Turner ML, Vandenberg CJ, Bouillet P, Metcalf D _et al_. Elevated Mcl-1 perturbs
lymphopoiesis, promotes transformation of hematopoietic stem/progenitor cells, and enhances drug resistance. _Blood_ 2010; 116: 3197–3207. Article CAS Google Scholar * Ogilvy S, Metcalf
D, Gibson L, Bath ML, Harris AW, Adams JM . Promoter elements of _vav_ drive transgene expression _in vivo_ throughout the hematopoietic compartment. _Blood_ 1999; 94: 1855–1863. CAS PubMed
Google Scholar * Egle A, Harris AW, Bath ML, O’Reilly L, Cory S . VavP-_Bcl2_ transgenic mice develop follicular lymphoma preceded by germinal center hyperplasia. _Blood_ 2004; 103:
2276–2283. Article CAS Google Scholar * Merino D, Khaw SL, Glaser SP, Anderson DJ, Belmont LD, Wong C _et al_. Bcl-2, Bcl-x(L), and Bcl-w are not equivalent targets of ABT-737 and
navitoclax (ABT-263) in lymphoid and leukemic cells. _Blood_ 2012; 119: 5807–5816. Article CAS Google Scholar * Pobezinsky LA, Angelov GS, Tai X, Jeurling S, Van Laethem F, Feigenbaum L
_et al_. Clonal deletion and the fate of autoreactive thymocytes that survive negative selection. _Nat Immunol_ 2012; 13: 569–578. Article CAS Google Scholar * Campbell KJ, Gray DH,
Anstee N, Strasser A, Cory S . Elevated Mcl-1 inhibits thymocyte apoptosis and alters thymic selection. _Cell Death Differ_ 2012; 19: 1962–1971. Article CAS Google Scholar * Strasser A,
Harris AW, Cory S . _Bcl-2_ transgene inhibits T cell death and perturbs thymic self-censorship. _Cell_ 1991; 67: 889–899. Article CAS Google Scholar * McCaughtry TM, Hogquist KA .
Central tolerance: what have we learned from mice? _Semin Immunopathol_ 2008; 30: 399–409. Article Google Scholar * Bouillet P, Purton JF, Godfrey DI, Zhang L-C, Coultas L, Puthalakath H
_et al_. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. _Nature_ 2002; 415: 922–926. Article CAS Google Scholar * Leeksma CH, Cohen JA .
Determination of the life of human blood platelets using labelled diisopropylfluorophosphanate. _Nature_ 1955; 175: 552–553. Article CAS Google Scholar * Ault KA, Knowles C . _In vivo_
biotinylation demonstrates that reticulated platelets are the youngest platelets in circulation. _Exp Hematol_ 1995; 23: 996–1001. CAS PubMed Google Scholar * Stewart DP, Koss B, Bathina
M, Perciavalle RM, Bisanz K, Opferman JT . Ubiquitin-independent degradation of antiapoptotic MCL-1. _Mol Cell Biol_ 2010; 30: 3099–3110. Article CAS Google Scholar * Kozuma Y, Yuki S,
Ninomiya H, Nagasawa T, Kojima H . Caspase activation is involved in early megakaryocyte differentiation but not in platelet production from megakaryocytes. _Leukemia_ 2009; 23: 1080–1086.
Article CAS Google Scholar * Schaefer BC, Schaefer ML, Kappler JW, Marrack P, Kedl RM . Observation of antigen-dependent CD8+ T-cell/ dendritic cell interactions _in vivo_. _Cell Immunol_
2001; 214: 110–122. Article CAS Google Scholar * White MJ, Kile BT . Apoptotic processes in megakaryocytes and platelets. _Semin Hematol_ 2010; 47: 227–234. Article CAS Google Scholar
* Mombaerts P, Iacomini J, Johnson RS, Herrup K, Tonegawa S, Papaioannou VE . RAG-1-deficient mice have no mature B and T lymphocytes. _Cell_ 1992; 68: 869–877. Article CAS Google
Scholar * Gray DH, Kupresanin F, Berzins SP, Herold MJ, O'Reilly LA, Bouillet P _et al_. The BH3-only proteins Bim and Puma cooperate to impose deletional tolerance of organ-specific
antigens. _Immunity_ 2012; 37: 451–462. Article CAS Google Scholar * Bouillet P, Metcalf D, Huang DCS, Tarlinton DM, Kay TWH, Köntgen F _et al_. Proapoptotic Bcl-2 relative Bim required
for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. _Science_ 1999; 286: 1735–1738. Article CAS Google Scholar * Merino D, Giam M, Hughes PD, Siggs OM,
Heger K, O'Reilly LA _et al_. The role of BH3-only protein Bim extends beyond inhibiting Bcl-2-like prosurvival proteins. _J Cell Biol_ 2009; 186: 355–362. Article CAS Google Scholar
* Ren D, Tu HC, Kim H, Wang GX, Bean GR, Takeuchi O _et al_. BID, BIM, and PUMA are essential for activation of the BAX- and BAK-dependent cell death program. _Science_ 2010; 330:
1390–1393. Article CAS Google Scholar * Czabotar PE, Lessene G, Strasser A, Adams JM . Control of apoptosis by the BCL-2 protein family and its therapeutic targeting. _Nat Rev Mol Cell
Biol_ 2013; 15: 49–63. Article Google Scholar * Zauli G, Vitale M, Falcieri E, Gibellini D, Bassini A, Celeghini C _et al_. _In vitro_ senescence and apoptotic cell death of human
megakaryocytes. _Blood_ 1997; 90: 2234–2243. CAS PubMed Google Scholar * Debrincat MA, Josefsson EC, James C, Henley KJ, Ellis S, Lebois M _et al_. Mcl-1 and Bcl-x(L) coordinately
regulate megakaryocyte survival. _Blood_ 2012; 119: 5850–5858. Article CAS Google Scholar * Kodama T, Hikita H, Kawaguchi T, Shigekawa M, Shimizu S, Hayashi Y _et al_. Mcl-1 and Bcl-xL
regulate Bak/Bax-dependent apoptosis of the megakaryocytic lineage at multistages. _Cell Death Differ_ 2012; 19: 1856–1869. Article CAS Google Scholar * Kodama T, Takehara T, Hikita H,
Shimizu S, Li W, Miyagi T _et al_. Thrombocytopenia exacerbates cholestasis-induced liver fibrosis in mice. _Gastroenterology_ 2010; 138: 2487–2498 2498 e1-7. Article CAS Google Scholar *
Kelly PN, White MJ, Goschnick MW, Fairfax KA, Tarlinton DM, Kinkel SA _et al_. Individual and overlapping roles of BH3-only proteins Bim and Bad in apoptosis of lymphocytes and platelets
and in suppression of thymic lymphoma development. _Cell Death Differ_ 2010; 17: 1655–1664. Article CAS Google Scholar * Mombaerts P, Iacomini J, Johnson RS, Herrup K, Tonegawa S,
Papaioannou VE . RAG-1-deficient mice have no mature B and T lymphocytes. _Cell_ 1992; 68: 869–877. Article CAS Google Scholar * Knudson CM, Tung KSK, Tourtellotte WG, Brown GAJ,
Korsmeyer SJ . Bax-deficient mice with lymphoid hyperplasia and male germ cell death. _Science_ 1995; 270: 96–99. Article CAS Google Scholar * Pezzella F, Tse AGD, Cordell JL, Pulford
KAF, Gatter KC, Mason DY . Expression of the _bcl_-2 oncogene protein is not specific for the 14;18 chromosomal translocation. _Am J Pathol_ 1990; 137: 225–232. CAS PubMed PubMed Central
Google Scholar Download references ACKNOWLEDGEMENTS We thank our colleagues Andreas Strasser, Warren Alexander, Daniel Gray and David Tarlinton for useful discussions; Giovanni Siciliano,
Jessica Mansheim, Kelly Trueman, Krystal Hughes, Sophie Allan, Marion Lebois and Katya Henley for excellent technical assistance; and A Strasser and D Tarlinton for reagents. This work was
supported by the NHMRC (Australia) program grants 461221; 1016647; NHMRC project grant 575535; NCI grant CA43540; Leukemia and Lymphoma Society Specialized Center for Research Grant 7015-02;
Australian Cancer Research Foundation (Centre for Therapeutic Target Drug Discovery); Fellowships from the Sylvia and Charles Viertel Foundation (BTK), Leukemia and Lymphoma Society (ECJ),
NHMRC Career Development Award (CJV); NHMRC/INSERM (CJ), EMBO (CJ, KJC), Human Frontier in Science Program (KJC) and infrastructure support to WEHI from the NHMRC Independent Research
Institute Infrastructure Support Scheme (IRISS) and the Victorian State Government Operational Infrastructure Support(OIS). AUTHOR CONTRIBUTIONS CJV and SC conceived the studies, planned
experiments, analysed data. BTK and ECJ provided advice and designed the platelet experiments. CJV, ECJ, KJC, CJ and KEL performed the experiments. CJV, SC, ECJ and BTK wrote the manuscript.
AUTHOR INFORMATION Author notes * K J Campbell Present address: Current address: Beatson Institute for Cancer Research, Switchback Road, Glasgow G61 1BD, UK, * C James Present address:
Current address: Laboratoire d'Hématologie du CHU de Bordeaux, Bordeaux, France; Université Bordeaux Segalen, Bordeaux, France; INSERM U1034, Pessac, Bordeaux, France, AUTHORS AND
AFFILIATIONS * Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade, Melbourne, Victoria 3052, Australia C J Vandenberg, E C Josefsson, K J Campbell, C James, K E Lawlor, B T
Kile & S Cory * Department of Medical Biology, University of Melbourne, Melbourne, Victoria 3010, Australia C J Vandenberg, E C Josefsson, K J Campbell, C James, K E Lawlor, B T Kile
& S Cory Authors * C J Vandenberg View author publications You can also search for this author inPubMed Google Scholar * E C Josefsson View author publications You can also search for
this author inPubMed Google Scholar * K J Campbell View author publications You can also search for this author inPubMed Google Scholar * C James View author publications You can also search
for this author inPubMed Google Scholar * K E Lawlor View author publications You can also search for this author inPubMed Google Scholar * B T Kile View author publications You can also
search for this author inPubMed Google Scholar * S Cory View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to S Cory.
ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no conflict of interest. ADDITIONAL INFORMATION Edited by C Borner Supplementary Information accompanies this paper on Cell Death
and Differentiation website SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION (PDF 1015 KB) RIGHTS AND PERMISSIONS This work is licensed under a Creative Commons
Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/ Reprints and permissions ABOUT THIS ARTICLE
CITE THIS ARTICLE Vandenberg, C., Josefsson, E., Campbell, K. _et al._ Loss of Bak enhances lymphocytosis but does not ameliorate thrombocytopaenia in _BCL-2_ transgenic mice. _Cell Death
Differ_ 21, 676–684 (2014). https://doi.org/10.1038/cdd.2013.201 Download citation * Received: 28 May 2013 * Revised: 11 December 2013 * Accepted: 16 December 2013 * Published: 24 January
2014 * Issue Date: May 2014 * DOI: https://doi.org/10.1038/cdd.2013.201 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link
Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * Bcl-2 * Mcl-1 * Bak *
apoptosis * thrombocytopaenia * haemopoiesis