Play all audios:
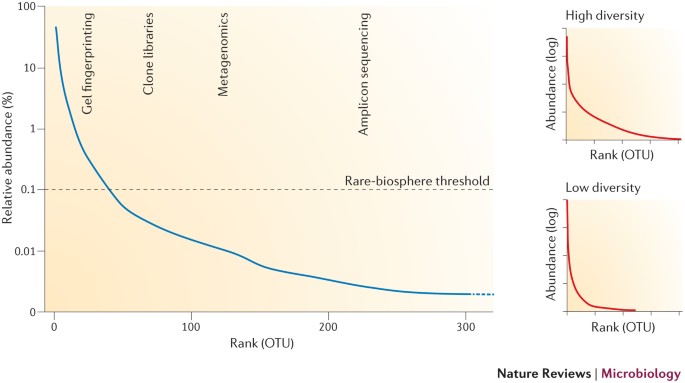
Microbial-community abundance distributions have a long 'tail' of low-abundance organisms, referred to as the rare biosphere, which often comprises the large majority of species.
Rare-biosphere microorganisms display specific and sometimes unique ecology and biogeography that can differ substantially from that of more abundant representatives.
The rare biosphere contributes to a persistent microbial seed bank, contrasting the influence of local microbial extinction and immigration.
Recruitment from the rare biosphere provides a broad reservoir of ecological function and resiliency (redundancy and flexibility).
Broad time-series studies with rich metadata will improve the study of rare-biosphere dynamics and conditionally rare taxa.
Study of the rare biosphere is prone to experimental artefacts (sequencing noise) and biological artefacts (dormancy and taphonomic gradients).
The majority of microbial diversity exists, at least transiently, in the rare biosphere.
Novel rare-biosphere members can be studied through cultivation, targeted phylogenetic mining and single-cell genomics.
The profound influence of microorganisms on human life and global biogeochemical cycles underlines the value of studying the biogeography of microorganisms, exploring microbial genomes and
expanding our understanding of most microbial species on Earth: that is, those present at low relative abundance. The detection and subsequent analysis of low-abundance microbial populations
— the 'rare biosphere' — have demonstrated the persistence, population dynamics, dispersion and predation of these microbial species. We discuss the ecology of rare microbial populations,
and highlight molecular and computational methods for targeting taxonomic 'blind spots' within the rare biosphere of complex microbial communities.
On page 6 the paper by Doxey, A. C. et al. entitled 'Aquatic metagenomes implicate Thaumarchaeota in global cobalamin production'. ISME J. 9, 461–471 (2015) should have been cited at the end
of the sentence ‘The recent discovery of thaumarchaeotal vitamin B12 production in aquatic environments also demonstrated winter archaeal abundance at station L4, in contrast to the low
relative abundance of related genes at other sampled time points’ instead of reference 75. We have added this new reference as Ref 181 in the reference list.
Woese, C. R. & Fox, G. E. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl Acad. Sci. USA 74, 5088–5090 (1977).
The authors appreciate comments provided by M. L. Sogin and B. J. Butler during the preparation of this manuscript. They acknowledge support from an Early Researcher Award (Government of
Ontario), the Canadian Institutes of Health Research (CIHR) and the Natural Sciences and Engineering Research Council of Canada (NSERC).
Department of Biology, University of Waterloo, Waterloo, N2L 3G1, Ontario, Canada
Although this has been an arbitrarily defined term, subject to variable abundance thresholds (for example,