Play all audios:
ABSTRACT Peptidoglycan hydrolases contribute to the generation of helical cell shape in _Campylobacter_ and _Helicobacter_ bacteria, while cytoskeletal or periskeletal proteins determine the
curved, vibrioid cell shape of _Caulobacter_ and _Vibrio_. Here, we identify a peptidoglycan hydrolase in the vibrioid-shaped predatory bacterium _Bdellovibrio bacteriovorus_ which invades
and replicates within the periplasm of Gram-negative prey bacteria. The protein, Bd1075, generates cell curvature in _B. bacteriovorus_ by exerting LD-carboxypeptidase activity upon the
predator cell wall as it grows inside spherical prey. Bd1075 localizes to the outer convex face of _B. bacteriovorus_; this asymmetric localization requires a nuclear transport factor 2-like
(NTF2) domain at the protein C-terminus. We solve the crystal structure of Bd1075, which is monomeric with key differences to other LD-carboxypeptidases. Rod-shaped _Δbd1075_ mutants invade
prey more slowly than curved wild-type predators and stretch invaded prey from within. We therefore propose that the vibrioid shape of _B. bacteriovorus_ contributes to predatory fitness.
SIMILAR CONTENT BEING VIEWED BY OTHERS A LYSOZYME WITH ALTERED SUBSTRATE SPECIFICITY FACILITATES PREY CELL EXIT BY THE PERIPLASMIC PREDATOR _BDELLOVIBRIO BACTERIOVORUS_ Article Open access
23 September 2020 EVIDENCE OF TWO DIFFERENTIALLY REGULATED ELONGASOMES IN _SALMONELLA_ Article Open access 09 September 2023 COORDINATED PEPTIDOGLYCAN SYNTHASES AND HYDROLASES STABILIZE THE
BACTERIAL CELL WALL Article Open access 02 September 2023 INTRODUCTION _Bdellovibrio bacteriovorus_ HD100 is a small, vibrioid-shaped predatory bacterium that invades and then replicates
within the periplasm of Gram-negative prey bacteria, forming a spherical structure called a prey bdelloplast1. _B. bacteriovorus_ has a broad prey range which includes multidrug-resistant
pathogens with variable outer membrane and cell wall chemistries, and the occurrence of genetic resistance to _B. bacteriovorus_ has never been observed in prey bacteria2,3. Predatory _B.
bacteriovorus_ can also successfully clear pathogen infections within a range of in vivo animal models4,5,6 and therefore has considerable and growing potential as a novel antibacterial
therapeutic. The predation process is critically dependent upon the modification of both predator and prey peptidoglycan (PG) cell walls to facilitate the dual bacterial encounter. PG forms
a complex macromolecular structure called a sacculus which surrounds the cytoplasmic membrane of nearly all bacteria, maintaining cell shape and providing protection against lysis due to
osmotic pressure fluctuations and large extracellular toxins7. Bacterial growth, cell division, and—importantly in this study—predation, occur through PG remodeling, which involves a
repertoire of predator-secreted modifying enzymes8,9,10,11. The predatory lifecycle of _B. bacteriovorus_ begins with attack-phase cells that swim12 or glide13 to encounter prey, then
recognize and attach to the prey outer membrane. An entry porthole in the prey cell wall is created, through which the predator traverses to enter the inner periplasmic compartment10.
Concurrently, two predator DD-endopeptidases are secreted into prey, cleaving crosslinks between prey PG peptide chains to sculpt rod-shaped prey cells into spherical bdelloplasts8. This
also reduces the frequency of sequential predator invasions, thus conferring exclusivity to the first-entering predator8. The porthole in the wall and outer membrane is then resealed and the
predator secretes hydrolytic enzymes including nucleases and proteases into the cytoplasm of the now-dead host, taking up the nutrient-rich degradative products14,15. Prey-derived and
de-novo-synthesized nucleotides are incorporated into the replicating genome copies of the predator which grows as an elongating multi-nucleoid filament inside the rounded but intact prey
until exhaustion of prey nutrients16. Synchronous septation of the predator filament yields progeny cells that secrete targeted PG hydrolytic enzymes to lyse the prey host and reinitiate the
predatory cycle11. PG hydrolases have an additional role generally in the determination of cell shape17, which has been particularly studied in the non-predatory, ε-proteobacteria
_Helicobacter pylori_18,19,20 and _Campylobacter jejuni_21,22,23, in whom multiple PG hydrolases collectively generate helical morphology. In contrast, bacterial vibrioid morphology is
generally determined by non-enzymatic cyto- or periskeletal proteins (well-studied in _Caulobacter crescentus_24,25 and _Vibrio cholerae_26,27). Despite the characterization of predator
enzymes that modify the prey PG, there have been very few studies concerning the cell wall PG architecture or vibrioid cell shape of predatory bacteria. Here, we investigate the mechanism by
which a curved, vibrioid predator is generated and ask whether there are evolutionary and functional connections between predator cell morphology and an efficient predatory lifestyle. Here,
we identify and characterize a predatory cell shape-determinant: Bd1075. Bd1075 is targeted to the outer convex cell face by its C-terminal nuclear transport factor 2-like (NTF2) domain,
where it exerts localized LD-carboxypeptidase (LD-CPase) activity upon the _B. bacteriovorus_ PG wall to generate curvature and the classical vibrio shape. We also show that the vibrioid
cell shape of _B. bacteriovorus_ predators facilitates rapid invasion into the periplasm of Gram-negative prey bacteria. RESULTS BD1075 GENERATES THE CURVATURE OF _B. BACTERIOVORUS_
PREDATORS The monocistronic _bd1075_ gene of the vibrioid-shaped _B. bacteriovorus_ Type strain HD100 encodes a 329 amino acid hypothetical protein with a predicted N-terminal sec signal
peptide28, suggestive of protein translocation into the periplasm or secretion from the cell (Supplementary Fig. 1). Bd1075 shares limited homology with Csd6 (identity: 24%, similarity: 38%)
and Pgp2 (identity: 25%, similarity: 40%) which are dimeric proteins important for the generation of helical cell shape in _H. pylori_18,29 and _C. jejuni_21, respectively. These
comparisons led us to hypothesize that Bd1075 could fulfill a role in the shape-determination of vibrioid predator _B. bacteriovorus_. Reverse-transcriptase PCR (RT-PCR) revealed that
_bd1075_ is constitutively transcribed throughout the predatory cycle, suggesting that the protein may have a role in _B. bacteriovorus_ rather than a secreted predatory function
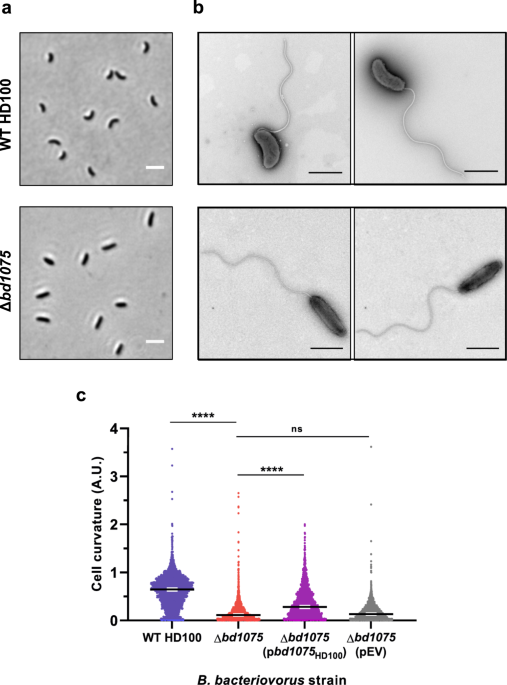
(Supplementary Fig. 2). A markerless deletion of _bd1075_ in the curved _B. bacteriovorus_ Type strain HD100 could still be cultured predatorily (phenotype differences further detailed
later) but ∆_bd1075_ mutant cells had a distinct straight rod-shaped morphology unlike the curved wild-type HD100 parent strain (Fig. 1a, b). Wild-type median curvature (0.64 A.U., 95% CI
[0.63, 0.66]) was significantly higher (_p_ < 0.0001; Fig. 1c) than the ∆_bd1075_ mutant (0.11 A.U., 95% CI [0.10, 0.12]). Plasmid-based complementation of ∆_bd1075_ with the wild-type
_bd1075_HD100 gene increased curvature relative to the ∆_bd1075_ mutant (Fig. 1c). These results indicate that Bd1075 has a role in generating the curvature of _B. bacteriovorus_. The
straight rod morphology of ∆_bd1075_ resembles the long-cultured laboratory strain _B. bacteriovorus_ 109J, which was isolated in the 1960s and is the only reported non-vibrioid strain of
_B. bacteriovorus_1. As _bd1075_ is conserved in all _B. bacteriovorus_ strains including 109J and appears to be a curvature-determinant, we queried why strain 109J is non-vibrioid. Despite
otherwise 100% sequence identity with _bd1075_HD100, _bd1075_109J contains an in-frame N-terminal truncation of 57 amino acids (Pro-18 to Tyr-74) (Supplementary Fig. 3a-c). RT-PCR confirmed
that _bd1075_109J is expressed in attack-phase cells and that the RNA transcript contains the predicted truncation (Supplementary Fig. 3d). To test whether the N-terminal truncation may
render the translated protein non-functional in curvature-determination, we cross-expressed _bd1075_109J in the HD100 ∆_bd1075_ mutant and found that this did not complement curvature
(Supplementary Fig. 4). In contrast, cross-expression of _bd1075_HD100 in wild-type 109J resulted in a median curvature of 0.29 A.U., 95% CI [0.26, 0.31], which is significantly higher than
the curvature of wild-type strain 109J (0.17 A.U., 95% CI [0.15, 0.18], _p_ < 0.0001; Supplementary Fig. 4). These results show that Bd1075 is a curvature-determinant of vibrioid _B.
bacteriovorus_ strains and that an inactivating mutation within the gene resulted in the lab-evolved strain 109J, which is unable to generate cell curvature. ROD-SHAPED PREDATORS INVADE PREY
MORE SLOWLY THAN THE CURVED WILD-TYPE As cell morphology can be phenotypically important in other bacteria, we asked whether the curved shape of _B. bacteriovorus_ could be advantageous to
the bacterial predator during its unique intraperiplasmic lifecycle. Comparison of the gross predation efficiency of wild-type and Δ_bd1075_ predators upon _E. coli_ prey in either liquid
culture, or on pre-grown _E. coli_ biofilms, did not reveal a significant difference (Supplementary Fig. 6 and Supplementary Fig. 7). However, these are laboratory conditions with readily
available prey and in which multiple important factors required to locate and navigate towards prey (e.g., predator chemotaxis and locomotion) are operational in bringing predators close to
the prey surface. We considered that predator morphology may fulfill an important role at the interface of single predator-prey encounters and therefore studied predation more closely at the
single-cell level using time-lapse microscopy to visualize individual predatory invasion events. _B. bacteriovorus_ HD100 wild-type or Δ_bd1075_ strains were mixed with _E. coli_ S17-1 and
placed under a microscope which captured images of specific fields of view every 1 min until the majority of _E. coli_ prey had been invaded. Hypothesizing that curvature may affect the
invasion of _B. bacteriovorus_ into prey, we measured two parameters: prey attachment time and prey entry time (Fig. 2a). Duration of prey attachment did not significantly differ (_p_ =
0.46) between the wild-type (median 28.5 min, 95% CI [28.0, 29.0]) and ∆_bd1075_ (median 29.5 min, 95% CI [29.0, 30.0]) (Fig. 2b); however, there was a significant difference (_p_ <
0.0001) between the rates at which the wild-type and ∆_bd1075_ predators entered prey: wild-type median 4.0 min (95% CI [4.0, 5.0]) versus ∆_bd1075_ median 6.0 min (95% CI [5.0, 6.0]),
respectively (Fig. 2c). Moreover, the longest wild-type entry was a single 7 min invasion, whereas 35.6% of ∆_bd1075_ entry invasions were ≥7 min, with the longest invasion lasting 14 min.
Complementation of ∆_bd1075_ by single-crossover reintroduction of the _bd1075_HD100 gene into the genome of ∆_bd1075_ restored entry time to wild-type levels (median 5.0 min, 95% CI [4.0,
5.0]; Fig. 2c). These data indicate that _B. bacteriovorus_ vibrioid morphology facilitates the traversal of predators across the prey cell envelope into the intraperiplasmic compartment of
the rounded prey cell. PREY BDELLOPLASTS ARE STRETCHED AND DEFORMED BY ROD-SHAPED PREDATORS Having observed that the non-vibrioid Δ_bd1075_ mutant was slower to enter the prey periplasm than
the curved wild-type, we next investigated the growth of Δ_bd1075_ within prey. During prey invasion, _B. bacteriovorus_ secretes two DD-endopeptidases, Bd0816 and Bd3459, into the prey
periplasm which hydrolyze the peptide bonds connecting chains of polysaccharide backbone8. The prey PG wall becomes more malleable and the cell rounds up into a spherical bdelloplast. We
hypothesized that growth of the straight rod-shaped Δ_bd1075_ within spherical bdelloplasts may be deleterious to the predatory niche, whereas curved wild-type cells may better ‘fit’ into
the curvature of the bdelloplast during growth and elongation. A C-terminal mCerulean3 fusion to the continuously-expressed cytoplasmic protein Bd00646,30 was introduced via single-crossover
recombination into both wild-type HD100 and Δ_bd1075_ to label the predator cytoplasm blue and allow visualization of _B. bacteriovorus_ within prey. Fluorescent _B. bacteriovorus_ strains
were mixed with _E. coli_ S17-1 pZMR100 and observed throughout the predatory cycle. Wild-type predators elongated as tightly curved filaments inside bdelloplasts (Fig. 3a); however, the
rod-shaped Δ_bd1075_ mutant—despite becoming more curved inside the spherical bdelloplast environment over time (Fig. 3b)—elongated as a less tightly curved filament and then septated to
give rod-shaped, non-vibrioid progeny cells (Fig. 3a). Strikingly, a subset of Δ_bd1075_ predator cells (~9.2%) appeared to stretch and deform the usually spherical prey bdelloplasts during
intrabacterial growth (Fig. 3c). Measuring the morphology of bdelloplasts containing a single _B. bacteriovorus_ predator between 1 h and 2.5 h after predator-prey mixing showed that from 1
to 2 h, the shape of prey bdelloplasts did not obviously differ between the two strains. However, at 2.5 h when _B. bacteriovorus_ cells are nearing maximal growth, the morphology of prey
bdelloplasts became markedly different. The median area of each bdelloplast containing a Δ_bd1075_ mutant predator (1.74 μm2, 95% CI [1.68, 1.79]) was significantly higher than bdelloplasts
containing wild-type predators (1.63 μm2, 95% CI [1.56, 1.73], _p_ < 0.01; Fig. 3d) and the median circularity was significantly lower (Δ_bd1075:_ 0.9925 A.U., 95% CI [0.9915, 0.9932];
wild-type: 0.9934 A.U., 95% CI [0.9931, 0.9951], _p_ < 0.05; Fig. 3e). Bdelloplasts containing Δ_bd1075_ predators were also significantly longer (median 1.55 μm, 95% CI [1.51, 1.58], _p_
< 0.01; Fig. 3f) than those containing curved wild-type predators (1.51 μm, 95% CI [1.47, 1.55]) but the median width did not significantly differ (Δ_bd1075:_ 1.37 μm, 95% CI [1.35,
1.39]; wild-type: 1.35 μm, 95% CI [1.32, 1.37], _p_ > 0.05; Fig. 3g), consistent with the visual appearance of stretched bdelloplasts. These findings suggest that rod-shaped mutant
predators can stretch and deform the spherical prey niche, in contrast to the curved wild-type. BD1075 EXERTS LD-CARBOXYPEPTIDASE ACTIVITY ON PG SACCULI IN VIVO AND IN VITRO Bd1075 contains
a predicted LD-transpeptidase (LDT) catalytic domain (Supplementary Fig. 1c, d); however, the LDT domains of related proteins Csd6 and Pgp2 function instead as LD-carboxypeptidases
(LD-CPases)18,21, which remove the terminal D-alanine from a PG tetrapeptide (consisting of L-Ala, D-Glu, _meso_-Dap, and D-Ala) to generate a tripeptide (consisting of L-Ala, D-Glu, and
_meso_-Dap)31. This highlighted the need to verify the catalytic activity (if any) of Bd1075. PG sacculi were therefore purified from _B. bacteriovorus_ strains and analyzed by HPLC to
determine their muropeptide composition and any changes to it caused by Bd1075. In contrast to curved wild-type HD100 sacculi, rod-shaped ∆_bd1075_ sacculi contained a greater proportion of
monomeric tetrapeptides (23.7 ± 0.8%) and crosslinked tetratetrapeptides (33.2 ± 0.7%) compared to the wild-type (9.6 ± 0.8% and 18.6 ± 0.6%, respectively), and a complete absence of
monomeric tripeptides and dimeric tetratripeptides (Fig. 4a, b and Table 1). This difference suggests that Bd1075 could cleave the C-terminal D-alanine of tetrapeptides to produce
tripeptides which terminate with _meso_-Dap. The complemented strain ∆_bd1075_ (p_bd1075_HD100) contained no monomeric tetrapeptides, 14.8 ± 1.2% tripeptides and 7.2 ± 0.5% dipeptides. These
data suggest that reintroduction of the wild-type _bd1075_HD100 gene resulted in over-complementation beyond wild-type as all monomeric tetrapeptides have been cleaved to tripeptides, with
some subsequently converted to dipeptides (Fig. 4c and Table 1). In comparison, the muropeptide profile of strain ∆_bd1075_ (p_bd1075_109J), in which curvature was not complemented
(Supplementary Fig. 4), did not differ from ∆_bd1075_, further confirming the non-functionality of truncated Bd1075109J as an LD-CPase (Supplementary Fig. 14a and Supplementary Table 1).
Wild-type 109J had a very similar muropeptide profile to the ∆_bd1075_ mutant—a complete absence of tripeptides and tetratripeptides and a high proportion of monomeric tetrapeptides (28.0 ±
4.3%) and dimeric tetratetrapeptides (33.8 ± 1.1%) (Supplementary Fig. 14b and Supplementary Table 1). Finally, the cross-complementation strain 109J (p_bd1075_HD100), with an increased
curvature compared to the lab-cultured wild-type strain 109J (Supplementary Fig. 4), contained a higher proportion of monomeric tripeptides (18.7 ± 3.0%) and dimeric tetratripeptides (26.2 ±
0.6%), and a reduction in monomeric tetrapeptides and dimeric tetratetrapeptides (3.0 ± 0.8% and 8.8 ± 0.8%, respectively) (Supplementary Fig. 14c and Supplementary Table 1). This
demonstrates that cross-expression of _bd1075_HD100 in wild-type 109J resulted in the enzymatic conversion of tetrapeptides to tripeptides and increased the curvature of this normally
non-vibrioid strain. To further validate the LD-CPase activity of Bd1075, an N-terminally His-tagged copy of _bd1075_HD100 was expressed in _E. coli_ BL21 and purified to near homogeneity by
Ni-NTA affinity chromatography and size-exclusion chromatography (Supplementary Fig. 15). The muropeptide profile of _B. bacteriovorus_ HD100 ∆_bd1075_ sacculi incubated with purified
Bd1075 enzyme revealed the complete conversion of both monomeric tetrapeptides to tripeptides and dimeric tetratetrapeptides to tetratripeptides (Fig. 4e). Bd1075 had identical enzymatic
activity on wild-type 109J straight rod sacculi and upon sacculi of wild-type _E. coli_ BW25113, showing that the enzyme can act on PG from different bacterial strains and species
(Supplementary Fig. 16b-c). These muropeptide data determine that Bd1075 has LD-CPase activity on PG both in vivo and in vitro, removing C-terminal D-alanine residues linked to the L-center
of _meso_-Dap to convert tetrapeptides to tripeptides. BD1075 STRUCTURE DETERMINATION The structure of mature Bd1075 protein was determined to 1.34 Å (Fig. 5 and Supplementary Table 3). The
Bd1075 structure contains two domains: the catalytic LD-CPase domain (aa 47–180) and a C-terminal nuclear transport factor 2 (NTF2)-like domain (aa 196–304) (Fig. 5a). Interestingly,
although there was a global agreement in the fold to Csd6 of _H. pylori_ and Pgp2 of _C. jejuni_ which also contain an NTF2 domain and an LD-CPase domain (the junction between the two at
residue 188 of Bd1075), there were significant differences in fold elements and local regions. These differences resulted in an inability to solve Bd1075 via molecular replacement,
necessitating the use of SAD phasing with co-crystallized halide ions. The Bd1075 protein is monomeric (the two molecules in the asymmetric unit contact one another by packing interactions
only), lacking the dimerization regions of the other characterized LD-CPase proteins; this was supported by size-exclusion data (Supplementary Fig. 18). We were able to trace residues 29–308
with the exception of a presumably flexible region (aa 82–91) which we term the active site ‘lid’. Differences to other LD-CPase structures are distributed throughout the fold (and in a
small shift in NTF2:LD-CPase juxtaposition) as demonstrated by RMSD values for the full-length/LD-CPase-alone/NTF2-alone of 2.5 / 2 / 1.8 Å for Csd6 and 2.8 / 1.9 / 2.2 Å for Pgp2. The large
values of 2.5/2.8 Å for full-length RMSD are in contrast with the agreement of 1.8 Å between Csd6 and Pgp2, hence Bd1075 is the structural outlier of the three proteins. Bd1075 has a
consensus active site, with the superfamily conserved catalytic triad consisting of C156, H141, and A142, each present in the expected (presumed active) orientations (Fig. 5b). A142 is often
a glycine residue in other LD-CPases but here it makes identical h-bonding contacts to H141 using backbone carbonyl atoms. Bd1075 active site pocket-forming residue V158 is a relative
anomaly as this position is arginine in most LD-transpeptidases or alanine in Csd6/Pgp2 and YafK-like enzymes shown recently to cleave the crosslink between PG and Braun’s lipoprotein32,33.
The termini of Bd1075 are very different to both Csd6 and Pgp2, replacing the N-terminal dimerization domain of those enzymes with a simple, shorter loop, and extending the C-terminus such
that the Bd1075 NTF2 domain finishes with a longer beta-strand (aa 295–306)—residues of which contribute to its binding pocket (Fig. 5d, e). The C-terminus has a further 21 residues that we
were unable to fit in this crystal form and which are predicted to be at least partly disordered. Most noteworthy within the core of the Bd1075 C-terminus was the presence of W303, which is
clearly located within the substrate-binding pocket of the NTF2 domain. W303 is highly conserved amongst _Bdellovibrio_ Bd1075 protein homologs but not found in other LD-CPase proteins (due
to their shorter NTF2 sequences which terminate at a position equivalent to Bd1075E302). The crystal packing of Bd1075 was such that a two amino acid loop of P107/K108 from one Bd1075
monomer packed into the NTF2 domain of the adjacent monomer. This loop is situated in an identical position to the bound glycerol of Csd6, postulated to be reminiscent of a substrate-like
interaction29. Conserved residues of the Bd1075 NTF2 domain binding pocket pack around this feature (yellow in Fig. 5c), the base of which is formed by Y274—an important residue in both our
monomeric structure and other dimeric LD-CPases. Having begun to probe the cellular localization of Bd1075 in _B. bacteriovorus_, we used this new structural information to aid in the
construction of fluorescently-tagged Bd1075 truncations and point mutants for enzyme localization tests. BD1075 IS TARGETED TO THE OUTER CONVEX CELL FACE BY ITS NTF2 DOMAIN To determine
whether Bd1075 is broadly active over all _B. bacteriovorus_ envelope PG or if an activity is specifically localized, a double-crossover markerless strain in which mCherry is C-terminally
fused to Bd1075 was constructed. Bd1075-mCherry localized to the outer convex face of _B. bacteriovorus_—both in free-swimming attack-phase cells (Fig. 6a) and throughout the predatory cycle
(Supplementary Fig. 19). The periplasmic localization of Bd1075 was tested via the construction of fluorescent fusions to mCitrine—a fluorophore incapable of periplasmic fluorescence.
Bd1075-mCitrine gave no fluorescent signal in contrast to a Bd0064-mCitrine cytoplasmic control, suggesting that Bd1075 localizes to the periplasm (Supplementary Fig. 20). Interestingly,
unlike polar-localized proteins, the mechanism by which bacterial proteins are targeted to one of the cell lateral sides has not been described. Intrigued and partly informed by the protein
structural data, we utilized the specifically targeted Bd1075 to investigate how bacterial proteins might be targeted to just one lateral side-wall. We hypothesized that, in addition to the
N-terminal signal peptide which targets the protein for translocation into the periplasm, Bd1075 may contain a second internal targeting sequence that directs the protein to just one lateral
periplasmic side-wall. We therefore constructed five different protein variants of Bd1075, which were each fused to mCherry at the protein C-terminus (Fig. 6b). These constructs were then
introduced into the _B. bacteriovorus_ HD100 curved wild-type strain to generate single-crossover strains containing two copies of _bd1075_: the original wild-type and the new mCherry
fusion. The correct and stable production of each fusion protein was confirmed by western blot analysis (Supplementary Fig. 21). The subcellular localization of each fusion protein was then
examined by epifluorescence microscopy. As expected, the full-length protein localized to the convex cell face (Fig. 6c) and we noted no morphological nor deleterious effects resulting from
the presence of two functional copies of _bd1075_. The LD-CPase domain contains three conserved catalytic triad residues: His-141, Ala-142, and Cys-156. Mutation of Cys-156 to alanine
(C156A) in full-length Bd1075 did not abrogate localization, indicating that LD-CPase activity is not involved in targeting (Fig. 6c). As the Bd1075 C-terminal NTF2 domain (aa 196–304) is
from a very broad protein superfamily34, it was not possible to predict a putative function for this domain but the structure with the P107/K108 loop bound over the NTF2 pocket (Fig. 5c)
suggested that this was a substrate interaction mimic and a means to destabilize putative PG substrate interactions. We tested a role for the NTF2 domain in protein targeting via three
mCherry fusions: (1) Full-length Bd1075 containing an NTF2 domain point mutation changing Tyr-274 (which forms the base of the substrate-binding pocket, Fig. 5c) to alanine: Y274A; (2)
Bd1075 truncated protein comprising residues 1-E302 (similar to the shorter Csd6 and Pgp2) which terminates 2 residues prior to the completion of the NTF2 domain at A304, omitting the highly
conserved W303 which the crystal structure had suggested to be a Bd1075-unique feature; and (3) Bd1075 truncated protein comprising residues 1-A304, completing the NTF2-like domain and
including W303. The E302 mutant, which contains a truncated and incomplete NTF2 domain, failed to localize to the outer curve (Fig. 6c), while the Y274A fusion was capable of only partial
localization (Fig. 6c), suggesting that this mutation in the NTF2 domain pocket may result in partial destabilization of the domain. In contrast, the A304 truncation mutant, which contains a
complete NTF2 domain (and _Bdellovibrio_-specific residue W303), was correctly targeted to the outer curve (Fig. 6c). These results strongly supported the hypothesis that the NTF2 domain is
involved in protein targeting to the convex cell face and that the _Bdellovibrio_-specific NTF2 extension, including W303, is important for this function. To investigate whether correct
protein targeting is absolutely required to generate curvature, the five mCherry fusion constructs were introduced into the rod-shaped Δ_bd1075_ mutant to generate single-crossover strains
expressing solely the mCherry-tagged copy of _bd1075_. mCherry fusions were again confirmed by western blot (Supplementary Fig. 21) and the subcellular localization and curvature of each
fusion strain were examined. The full-length protein localized to the outer curve and completely complemented the curvature of Δ_bd1075_ (Fig. 6d, e). The LD-CPase catalytic domain point
mutant C156A did not restore curvature and was diffuse throughout the rod-shaped cell (Fig. 6d, e). Critically, neither of the NTF2 domain mutants Y274A nor E302 were correctly targeted nor
could either protein complement the curvature of Δ_bd1075_ (Fig. 6d, e), highlighting the importance of the NTF2 domain and residue W303. Together, these targeting data reveal that the NTF2
domain is responsible for targeting Bd1075 to the outer curve of the bacterial periplasm and that this specific localization is required to generate cell curvature. DISCUSSION In this work,
we elucidate the vibrioid cell shape-determinant of predatory _Bdellovibrio bacteriovorus_ bacteria and show that vibrioid morphology facilitates rapid prey invasion. These findings
contribute to fundamental knowledge of bacterial cell shape and deepen our understanding of the predatory process, which may assist the application of predatory bacteria as a therapeutic. In
vibrioid bacteria, intermediate filament-like (IF-like) cyto- or periskeletal elements frequently determine cell shape24,26. IF-like proteins often contain coiled-coil rich protein (Ccrp)
domains; however, deletion of the sole _B. bacteriovorus_ Ccrp protein was previously found not to affect vibrioid cell shape35. Here, we discover that the vibrioid curvature of _B.
bacteriovorus_ is instead determined by a PG cell wall hydrolase: Bd1075. Refining the initial prediction of an LDT domain, we show via sacculus studies that Bd1075 functions as an LD-CPase,
cleaving both crosslinked and uncross linked tetrapeptides to tripeptides in the predator PG cell wall (Fig. 4 and Table 1). This enzymatic activity was also observed for LD-CPase helical
shape-determining proteins Csd6 (_H. pylori_) and Pgp2 (_C. jejuni_), highlighting the importance of biochemical validation of predicted LDT domains. It is interesting to note that, while
plasmid-based complementation of Δ_bd1075_ with the wild-type _bd1075_HD100 gene resulted in partial complementation of cell curvature (Fig. 1c), the same strain showed ‘over-complemented’
enzymatic activity on PG, with complete conversion of tetrapeptides to tripeptides and subsequent cleavage to dipeptides (Fig. 4c and Table 1). A similar phenomenon was observed for the
LD-CPase protein Csd6 and was caused by a 2.1× increase in protein production compared to the wild-type18, whereby an excess of enzymatic activity resulted in ‘under-complementation’ of
curvature. This suggests that an incorrect level of enzymatic activity leads to dysregulation of the system in general and the same is likely true here for Bd1075. The overall peptide
crosslinkage was slightly higher in cells lacking _bd1075_ (64.4% compared to 61.1% in wild-type, Table 1). The reason for this effect is enigmatic but we hypothesize that a localized
reduction in PG crosslinkage in wild-type cells could soften one lateral side-wall of the PG sacculus such that the cell bulges slightly and becomes deformed by internal cellular turgor
pressure which pushes outwards to generate an outer convex curve that may be fixed by subsequent enzyme activity. A similar mechanism has been proposed to explain cell shape in _H. pylori_
and _C. jejuni_19,20,21,36. _B. bacteriovorus_ vibrioid curvature is widely conserved within this group of invasive predators with the exception of the rod-shaped and long-cultured
laboratory strain _B. bacteriovorus_ 109J. Originally named _B. bacteriovorus_ 109, the strain was redesignated as 109J following the observation that, in one research laboratory, predator
cells had transitioned from a curved to non-vibrioid shape37. It is possible that long-term laboratory culture conditions in which prey are highly abundant may have removed the selection
pressure for vibrioid morphology, resulting in the lab-evolved 57-residue deletion that we detect to have inactivated Bd1075109J. Bd1075109J was not catalytically active upon PG
(Supplementary Fig. 14a, Supplementary Fig. 16b, and Supplementary Table 1) and therefore could not complement curvature when cross-expressed in HD100 Δ_bd1075_ (Supplementary Fig. 4).
Despite retaining domains for LD-CPase catalytic residues and targeting capability, the 57-residue N-terminal truncation could severely disrupt Bd1075109J protein folding and therefore
function. The mutation most likely occurred via homologous recombination between 8 bp repeats which flank the deleted region (Supplementary Fig. 3b); this has been observed in other predator
genes38. Bacterial morphology is evolutionarily conserved and known to confer selective advantages to different bacterial lifestyles39,40. The helical morphology of _H. pylori_, for
example, facilitates efficient bacterial motility through the gastric mucosa to allow pathogenic colonization of the gastrointestinal tract19. Moreover, rod-shaped mutants of helical _C.
jejuni_ are deficient or have reduced fitness in a model of chick colonization21,22. In _V. cholerae_, vibrioid cell curvature (which is generated via a different non-enzymatic mechanism)
increases motility through dense soft-agar matrices and promotes the colonization of _V. cholerae_ in motility-dependent mice and rat infection models26. Here, we propose that _B.
bacteriovorus_ cell curvature facilitates efficient predatory invasion into Gram-negative prey bacteria. Straight rod-shaped _B. bacteriovorus_ Δ_bd1075_ predators invade prey significantly
more slowly than curved wild-type predators (Fig. 2c). During prey invasion, _B. bacteriovorus_ must presumably overcome opposing physical forces exerted upon itself by the turgid prey
membrane and cell wall. Curved predators may distribute opposing forces as a glancing blow along the predator cell body, facilitating an efficient, curved trajectory into rounded prey, in
contrast to force-intensive head-on invasions by rod-shaped predators. Unlike wild-type curved _B. bacteriovorus_, non-vibrioid Δ_bd1075_ predators stretch and deform the rounded prey
bdelloplast (Fig. 3c-e). Intriguingly, non-vibrioid Δ_bd1075_ predators become gradually curved during elongation inside spherical prey bdelloplasts, despite the absence of _bd1075_ (Fig.
3a, b). The Δ_bd1075_ predator does not curve as tightly as the wild-type, however, and released progeny cells are not curved but rod-shaped, indicating that adoption of an intracellular
curvature is temporary in this mutant (Fig. 3a). Various studies have shown that bacteria can sense and adapt their shape in response to different physiological topologies and adjust their
internal physiology accordingly41,42,43. For example, _E. coli_ cells that are subjected to mechanical strain within a confined environment can temporarily adopt the shape of that
environment but then recover wild-type morphology upon release from confinement44,45,46. The mechanisms of shape-sensing and restoration of wild-type morphology are not yet fully understood,
however47. Here, it is possible that in curved wild-type predators, Bd1075 may sense the temporary curvature imposed by the bdelloplast and enzymatically act on the _B. bacteriovorus_ PG
wall to tighten and permanently fix this curvature. This avoids potential damage to the replicative bdelloplast niche, while also preparing curved and invasively streamlined progeny
predators for prey exit. Fluorescently-tagged Bd1075 specifically localizes to the outer convex face of vibrioid _B. bacteriovorus_ cells (Fig. 6a). Prior to this study, most cell shape
enzymes (for which localization is known) were found at the inner concave face of bacteria. These include the _C. crescentus_ cytoskeletal polymer crescentin24 and _Vibrio cholera_e CrvA and
CrvB which form periskeletal polymers26,27. Until the discovery of Bd1075, only the _H. pylori_ bactofilin CcmA had been identified at the outer convex face, where it occupies the
cytoplasm36, unlike Bd1075 which is periplasmic (Supplementary Fig. 20). To our knowledge, there is currently no known mechanistic basis for asymmetric bacterial protein targeting to one
lateral side-wall of the cell. Intrigued by this and guided by the phenotyping of Bd1075 protein truncations and point mutations, we discover that the extended C-terminal NTF2 domain
(including the unique pocket residue W303) targets the protein to the convex face and is necessary to generate cell curvature (Fig. 6c-e). NTF2 is a nuclear envelope protein that transports
molecules into eukaryotic nuclei48. The NTF2-like domain superfamily, however, comprises hundreds of thousands of proteins spanning the 3 domains of life and is associated with over 200
biological pathways, suggestive of divergent evolution34,49. The NTF2 domain of _C. jejuni_ Pgp2 was recently found by NMR studies to bind a variety of PG fragments, with specific secondary
structure features shifting upon complexation50. The general agreement of some of the structures of monomeric Bd1075 with dimeric Pgp2 is suggestive that Bd1075 binds PG. The
hydrogen-bond-rich nature of the P107/K108 loop we observe in our crystal structure could be indicative of such interaction by mimicking part of a PG muropeptide. Since the Pgp2 study
invoked an induced fit on binding PG, it is particularly interesting that the third helix of the Bd1075 NTF2 domain (nH3, aa 233–248) and associated loop (labeled ‘lip’, aa 226–232) are
shifted in relation to both Pgp2 and Csd6 (Fig. 5d). This helix was the major NTF2 feature that was reported to shift upon Pgp2-PG binding50. We postulate that its position has been modified
in our Bd1075 structure by sidechains contacting the P107/K108 loop and may thus represent a ‘bound’ state. There is also the potential for NTF2:LD-CPase domain crosstalk, given that a
small LD-CPase domain helix (aa 125–132) shifts in response to NTF2 alterations50, and in Bd1075 appears to influence the disorder of the adjacent active site lid domain. Since Bd1075
localizes to one lateral side-wall, one could theorize that the PG or outer membrane properties of that particular side-wall must be uniquely different from the other. One possibility is
that the NTF2 domain recognizes a modification or substrate which is more abundant at this side-wall. Alternatively, the NTF2 domain could sense the temporary physical curvature imposed by
growth inside the spherical bdelloplast (‘curvature-templating’) and direct Bd1075 to this curved cell face. Bd1075 may then exert LD-CPase activity on the PG wall, initiating the fixation
of _B. bacteriovorus_ curvature. An alternative idea, which should not be excluded, is that the existing localization of Bd1075 at the convex face drives the targeting of newly synthesized
Bd1075 to the convex face via self-interactions involving the NTF2 domain. These questions—which would present a significant investigative challenge beyond the scope of this study—could help
to explain how bacteria both sense and respond to different topological space environments. Collectively, these findings advance our understanding of factors affecting the physiology of
therapeutically promising predatory bacteria and provide mechanistic insights into the evolutionary importance of bacterial cell morphology. METHODS BACTERIAL STRAINS AND CULTURE
_Bdellovibrio bacteriovorus_ strains were predatorily cultured in liquid Ca/HEPES buffer (5.94 g/l HEPES free acid, 0.284 g/l calcium chloride dihydrate, pH 7.6) and on solid YPSC overlays,
both containing _Escherichia coli_ S17-1 prey as described previously51. Where appropriate, kanamycin or gentamicin was added to growth media at concentrations of 50 or 5 µg ml−1,
respectively. PLASMID AND STRAIN CONSTRUCTION Primers and plasmids used to construct strains used in this study are detailed in Supplementary Table 4 and Supplementary Table 5. Strains are
listed in Supplementary Table 6. Bd1075 protein residue numbering was updated to correct for a previously mis-annotated start codon; the probable true start begins 2 codons downstream from
that originally predicted (Supplementary Fig. 1b). To construct a markerless deletion of _bd1075_, 1 kb of upstream and downstream DNA were cloned into the suicide vector pK18_mobsacB_ by
Gibson assembly52 using the NEBuilder HiFi DNA Assembly Cloning Kit (New England Biolabs). The vector was introduced into _B. bacteriovorus_ via an _E. coli_ S17-1 conjugal donor strain and
a double-crossover deletion mutant was isolated by sucrose suicide counter-selection as described previously8,53. Deletion of _bd1075_ was verified by RT-PCR, Sanger sequencing, and
whole-genome sequencing. Strains for complementation tests were constructed by inserting the _bd1075_ gene from either strain HD100 or strain 109J (gene ID: EP01_15440) plus the respective
native gene promoter into the vector pMQBAD, a derivative of pMQ414 which expresses the fluorescent tdTomato protein and is capable of autonomous replication in _B. bacteriovorus_54.
Constructs for complementation tests were verified by Sanger sequencing, conjugated into _B. bacteriovorus_ and maintained under a gentamicin selection pressure. To initially
fluorescently-tag Bd1075, the stop codon of _bd1075_ was replaced with the _mCherry_ coding sequence to generate an in-frame C-terminal fusion terminating at the stop codon of the mCherry
protein. The fluorescent strain was constructed and verified analogously to the ∆_bd1075_ mutant to generate a markerless double-crossover Bd1075-mCherry strain. For a more extensive
analysis of domain functions, single-crossover mCherry fusions to either full-length or truncated versions of Bd1075 were made. These were constructed by cloning 1 kb of upstream DNA, the
DNA encoding each desired Bd1075 domain-test (minus the stop codon), and the _mCherry_ sequence into pK18_mobsacB_. Point mutations in such constructs were generated using Q5® Site-Directed
Mutagenesis (New England Biolabs). The Bd1075-mCitrine fusion (Supplementary Fig. 20) was constructed in an analogous manner but replaced the _mCherry_ sequence with _mCitrine_. Each
construct was then conjugated into _B. bacteriovorus_ to generate single-crossover merodiploid strains which were confirmed by Sanger sequencing and maintained under a kanamycin selection
pressure. PROTEIN EXPRESSION AND PURIFICATION The _bd1075_ gene from _B. bacteriovorus_ HD100, minus the signal peptide and stop codon, was cloned into the vector pET41 in-frame with a
C-terminal histidine tag and transformed into _E. coli_ BL21. _E. coli_ BL21 was cultured in TB media and incubated at 37 °C with orbital shaking at 200 rpm to an OD600 of 0.6–0.8. _bd1075_
expression was then induced with 0.5 mM IPTG at 18 °C for 16 h. Bd1075 was purified to near homogeneity using Ni-NTA affinity and size-exclusion chromatography, then dialyzed into either
buffer containing 10 mM Na Citrate pH 6.0, 30 mM KCl, and 2% w/v glycerol (activity assays) or buffer containing 20 mM Na Citrate pH 6.0, 200 mM NaCl and 2 mM β-mercaptoethanol (structure
studies). MUROPEPTIDE ANALYSIS To culture _B. bacteriovorus_ for sacculi isolation, 1 l of each _B. bacteriovorus_ strain was grown on either _E. coli_ S17-1 or _E. coli_ S17-1 pUC19
(gentamicin-resistant prey for complementation test strains containing the pMQBAD plasmid). Following complete predatory lysis of _E. coli_ prey during a 24 h incubation at 29 °C with
orbital shaking at 200 rpm, _B. bacteriovorus_ attack-phase cultures were passed through a 0.45 µm filter to remove any remaining prey debris. To culture _E. coli_ BW25113 for sacculi
isolation, _E. coli_ was grown at 37 °C for 16 h with orbital shaking at 200 rpm. Cultured _B. bacteriovorus_ or _E. coli_ cells were then centrifuged at 15,000 × _g_ for 30 min at 4 °C,
resuspended in 6 ml of ice-cold PBS, and then boiled in 6 ml of 8% SDS for 30 min to lyse the cells and liberate sacculi. Peptidoglycan was purified from the cell lysates, then muropeptides
in the supernatant were reduced with sodium borohydride, and HPLC analysis performed as previously described55. The major muropeptides (peak no. 1–7) were assigned by comparing the elution
times to those of well-characterized corresponding muropeptides from _E. coli_ and other Gram-negative bacteria in the published literature55,56. The identity of the muropeptides with lower
abundance and late elution times (peak no. 8–19) was confirmed by collecting and analyzing peaks from the WT HD100 chromatogram by mass spectrometry as described57. Mass spectrometry data
are reported in Supplementary Table 2. To test the in vitro activity of Bd1075HD100, sacculi from either _B. bacteriovorus_ HD100 ∆_bd1075_, _B. bacteriovorus_ wild-type 109J or _E. coli_
BW25113 were incubated with 10 µM of _B. bacteriovorus_ Bd1075HD100 in 50 mM Tris-HCl, 50 mM NaCl, pH 7.0 for 16 h at 37 °C on a thermomixer at 900 rpm. The control sample received no
enzyme. To stop Bd1075 activity, the samples were boiled at 100 °C for 10 min and an equal volume of 80 mM sodium phosphate, pH 4.8, was added. The samples were incubated with 10 µg of
cellosyl (Hoechst, Frankfurt am Main, Germany) for a further 16 h at 37 °C on a thermomixer at 900 rpm, boiled for 10 min and centrifuged at room temperature for 15 min at 16,000 × _g_.
Muropeptides present in the supernatant were reduced with sodium borohydride, analyzed by HPLC55 and identified as described above. STRUCTURE DETERMINATION Purified Bd1075 at ~25 mg/ml was
used for screening. Crystals were grown at 18 °C using the sitting drop technique in 4 μl drops composed of equal volumes of protein and reservoir solution. Bd1075 crystals were obtained in
the BCS™ screen (Molecular Dimensions) in condition 2–44, comprising 0.1 M Tris pH 7.8, 0.1 M KSCN, 0.1 M NaBr and 25% PEG Smear broad range. Crystals were cryo-protected in mother liquor
supplemented with 25% (v/v) ethylene glycol and flash cooled in liquid nitrogen. Diffraction data were collected at the Diamond Light Source in Oxford, UK (Supplementary Table 3). Data
reduction and processing were completed using XDS58 and the xia2 suite59. Bd1075 phasing was achieved using a merged SAD dataset (9000 frames, 0.1° oscillations) collected at a wavelength of
0.91 Å corresponding to the bromide anomalous scattering peak. The collected data were input into CCP4 online CRANK260, which located six bromide sites with an initial FOM of 0.14, followed
by iterative cycles of building and model-based phasing improvement. The obtained model was further built and modified using COOT61, with cycles of refinement in PHENIX62. PHASE-CONTRAST
AND EPIFLUORESCENCE MICROSCOPY _B. bacteriovorus_ cells were immobilized on a thin 1% Ca/HEPES buffer agarose pad and visualized under a Nikon Ti-E inverted epifluorescence microscope
equipped with a Plan Apo ×100 Ph3 oil objective lens (NA: 1.45), an mCherry filter (excitation: 555 nm, emission: 620/60 nm), a CFP filter for mCerulean3 (excitation: 440 nm, emission:
470–490 nm) and a YFP filter for mCitrine (excitation: 508 nm, emission 540/25 nm). Images were acquired on an Andor Neo sCMOS camera with Nikon NIS software. IMAGE ANALYSIS Images were
analyzed using the Fiji distribution of ImageJ63 and minimally processed using the sharpen and smooth tools, with adjustments to brightness and contrast. The MicrobeJ plug-in for ImageJ64
was used to measure cell morphologies and detect fluorescent signal (foci). All images were manually inspected to ensure cells had been correctly detected before measurements were acquired.
_B. bacteriovorus_ attack-phase cells were generally identified by the parameters of area: 0.2–1.5 µm2, length: 0.5–2.5 µm, width: 0.2–0.8 µm, and circularity: 0–0.9 A.U. To measure the
curvature of _B. bacteriovorus_ cells, MicrobeJ determines this as ‘the reciprocal of the radius of curvature measured between the endpoints and the center of the medial axis of the cell’
and it is described in arbitrary units (A.U.) (Supplementary Fig. 22). The fluorescent Bd1075-mCherry signal within attack-phase _B. bacteriovorus_ cells was detected with the most
appropriate MicrobeJ method (foci) using default maxima settings and an association with parent bacteria with a tolerance of 0.1 µm. Using this method, heatmaps could be generated to show
the distribution of fluorescence signals across large numbers of cells. For the detection of Bd1075-mCherry fluorescent fusions in a curved wild-type genetic background, _B. bacteriovorus_
cells were defined with the same parameters, but excluded cells with a curvature of <0.6 so as to only measure localization in cells with a definitively curved shape. Curvature parameters
were set to 0-max to allow measurements of curvature for fluorescent fusions expressed in a non-curved ∆_bd1075_ genetic background. To analyze prey bdelloplast morphology, bdelloplasts
were generally identified by the parameters of area: 1.0-max µm2, length: 0.5-max µm, width: 0.5-max µm, and circularity: 0.8–1.0 A.U. Circularity is defined in MicrobeJ as ‘4πr ×
area/perimeter2’ with a value of 1.0 indicating a perfect circle (Supplementary Fig. 22). Bdelloplasts with a circularity of <0.96 were classified as non-circular and stretched by
Δ_bd1075_ based on extensive visual observations and the fact that 0% of WT bdelloplasts had a circularity of <0.96 A.U. _B. bacteriovorus_ cells were detected in the maxima channel with
the bacteria method using default maxima settings and an association with parent bacteria with a tolerance of 0.1 µm. Only the morphologies of prey bdelloplasts which contained a single _B.
bacteriovorus_ predator were measured. ELECTRON MICROSCOPY _B. bacteriovorus_ cells were cultured for 24 h, then concentrated by microcentrifugation at 5000 × _g_ for 10 min followed by
careful resuspension in 1 ml of Ca/HEPES. _B. bacteriovorus_ cells were applied to glow-discharged Formvar/Carbon-coated 200-mesh copper grids (EM Resolutions), stained with 0.5% uranyl
acetate for 1 min, then de-stained with Tris-EDTA pH 7.6 for 30 s. Samples were imaged under a FEI Tecnai G2 12 Biotwin transmission electron microscope at 100 kV. TIME-LAPSE MICROSCOPY For
time-lapse microscopy, 1 ml cultures of attack-phase _B. bacteriovorus_ and 50 µl of stationary-phase _E. coli_ S17-1 were microcentrifuged separately at 17,000 × _g_ for 2 min, then
resuspended in 50 µl of Ca/HEPES. Predators and prey were then mixed together and immediately transferred to a thin 0.3% Ca/HEPES agarose pad. Cells were visualized under a Nikon Eclipse
E600 upright microscope equipped with a ×100 oil objective lens (NA: 1.25) and a Prior Scientific H101A XYZ stage, which allowed six specific fields of view to be revisited over the
time-lapse sequence. Image frames were captured every 1 min for at least 2 h on a Hammamatsu Orca ER Camera with Simple PCI software. Time-lapse videos of _B. bacteriovorus_ attaching to and
entering _E. coli_ prey were analyzed in Simple PCI software. Attachment time was measured by counting the number of frames (1 frame = 1 min) between initial irreversible predator
attachment to prey and the first indication of the predator moving into prey. Entry time was measured by counting the number of frames between the first indication of the predator moving
into prey and the predator residing completely inside the bdelloplast. REVERSE-TRANSCRIPTASE PCR To monitor the expression of _bd1075_ across the predatory cycle, RNA template was isolated
from different time points during a synchronous predation of _B. bacteriovorus_ HD100 on _E. coli_ S17-1 using an SV Total RNA Isolation System kit (Promega) as previously described65. RNA
quality was verified on an Agilent Bioanalyzer using an Agilent RNA 6000 Nano Kit. RT-PCR was carried out using the QIAGEN OneStep RT-PCR kit with the following thermocycling parameters: 50
°C for 30 min, 94 °C for 15 min, followed by 30 cycles of 94 °C for 1 min, 50 °C for 1 min, and 72 °C for 1 min, and a final step of 72 °C for 10 min. Samples were run on a 2% agarose gel at
100 V for 30 min. Full, uncropped gel images are available in the Source Data file. _B. BACTERIOVORUS_ PREDATION ON _E. COLI_ IN LIQUID CULTURE The predation efficiency of _B.
bacteriovorus_ strains in liquid culture was measured using a luminescent prey assay developed by Lambert et al., 200366. _B. bacteriovorus_ strains were cultured in Ca/HEPES buffer at 29 °C
with orbital shaking at 200 rpm for 24 h. Wild-type _B. bacteriovorus_ HD100 and Δ_bd1075_ predator strains were then matched by total protein content which was determined by Lowry assay67.
Matched _B. bacteriovorus_ strains were enumerated on overlay plates upon which plaques emerged after 5–7 days. Luminescent _E. coli_ S17-1 was cultured in YT broth at 37 °C with orbital
shaking at 200 rpm for 16 h. The stationary-phase _E. coli_ was then adjusted to OD600 0.2 in a 1:1 mixture of Ca/HEPES and PY medium and 200 μl were aliquoted into each well of a 96-well
microtiter optiplate. _E. coli_ prey were enumerated on YT agar plates at 37 °C for 16 h and typically had a concentration of 107 CFU ml−1. One milliliter samples of _B. bacteriovorus_ were
heat-killed by incubation at 105 °C for 5 min. To create a predator dilution series, _B. bacteriovorus_ live cell volumes of 0, 1, 2, 4, 8, 16, 32, and 64 μl were aliquoted into each well
containing _E. coli_ prey and made up to a total of 64 μl with heat-killed predator cells. Microtiter plates were covered with a Breathe-Easy® membrane and reduction in _E. coli_
luminescence was measured at 30 min intervals in a BMG FluoStar microplate reader maintained at 29 °C with double orbital shaking at 200 rpm. To compare the rates of _E. coli_ death by
predation, the area under each luminescence curve was measured and then normalized to the maximum luminescence for each dilution. Data were analyzed in BMG LABTECH MARS data analysis
software. _B. BACTERIOVORUS_ PREDATION ON _E. COLI_ BIOFILMS The predation efficiency of _B. bacteriovorus_ strains on prey biofilms was measured using an assay adapted from Lambert &
Sockett, 201353 and Medina et al., 200868. _E. coli_ S17-1 was cultured in YT broth at 37 °C with orbital shaking at 200 rpm for 16 h and then 200 μl of stationary-phase _E. coli_, adjusted
to OD600 0.1 in fresh YT, were aliquoted into each well of a 96-well PVC microtiter plate and incubated in a 29 °C static incubator for 24 h to produce a prey biofilm. _E. coli_ prey were
enumerated on YT agar plates at 37 °C for 16 h and typically had a concentration of 107 CFU ml−1. _B. bacteriovorus_ WT HD100 and Δ_bd1075_ were concurrently cultured at 29 °C with orbital
shaking at 200 rpm for 24 h. Following complete prey lysis, _B. bacteriovorus_ predators were filtered through a 0.45 μm membrane to remove residual prey. Predator strains were then matched
by total protein and plaques were enumerated on double-layer overlay plates after 5–7 days. The _E. coli_ biofilm-coated plate was removed from the incubator and carefully washed three times
with Ca/HEPES buffer to remove residual planktonic cells. _B. bacteriovorus_ 200 μl aliquots of neat (100), 10−1, 10−2, and 10−3 dilutions in Ca/HEPES buffer were then added to the _E.
coli_ biofilm plate. _B. bacteriovorus_ filtered through a 0.22 μm membrane served as a no-predator control. The 96-well plate containing predators and prey was incubated for a further 24 h
in a static 29 °C incubator. The plate was then stained with 1% crystal violet for 15 min, washed with sterile distilled water, and de-stained with 33% acetic acid for 15 min. Plate contents
were mixed carefully and then absorbance readings were acquired at OD600 to quantify the amount of _E. coli_ biofilm remaining. WESTERN BLOT ANALYSIS OF BD1075-MCHERRY FUSIONS _B.
bacteriovorus_ Bd1075-mCherry fusion strains were cultured for 24 h until prey were completely lysed, filtered through a 0.45 μm membrane to remove residual prey debris, and then centrifuged
at 5525 × _g_ for 20 min. The cell pellet was resuspended in 100 μl of 4× PAGE loading buffer containing β-mercaptoethanol, boiled for 5 min and then loaded onto a 4-20 % SDS-PAGE gel with
either a MagicMark™ XP Western protein standard ladder (for blotting gel) or a SeeBlue™ Plus2 Pre-stained Protein Standard ladder (for loading control gel). Gels were run at 150 V for 1 h.
Loading control gels were then stained and de-stained with QuickBlue Protein Stain (LubioScience). Blotting gels were transferred to a nitrocellulose membrane for 2 h at 25 V. Western blots
used an anti-mCherry primary antibody (Invitrogen, product no: PA5-34974, diluted 1:4000) and a WesternBreeze™ Chemilumincescent kit according to the manufacturer’s instructions. Images were
captured by exposure to X-ray film. STATISTICAL ANALYSIS Statistical analysis was performed in Prism 8.0 (GraphPad). Data were first tested for normality and then analyzed using the
appropriate statistical test. The number of biological repeats carried out, n values for cell numbers, and the statistical test applied to the dataset are described in each figure legend.
REPORTING SUMMARY Further information on research design is available in the Nature Research Reporting Summary linked to this article. DATA AVAILABILITY The Bd1075 crystal structure data
generated in this study have been deposited in the PDB database under the accession code 7O21. Source data are provided with this paper. REFERENCES * Stolp, H. & Starr, M. P.
_Bdellovibrio bacteriovorus_ gen. et sp. n., a predatory, ectoparasitic, and bacteriolytic microorganism. _Antonie Van. Leeuwenhoek_ 29, 217–248 (1963). Article CAS PubMed Google Scholar
* Dashiff, A., Junka, R. A., Libera, M. & Kadouri, D. E. Predation of human pathogens by the predatory bacteria _Micavibrio aeruginosavorus_ and _Bdellovibrio bacteriovorus_. _J. Appl.
Microbiol._ 110, 431–444 (2011). Article CAS PubMed Google Scholar * Kadouri, D. E., To, K., Shanks, R. M. & Doi, Y. Predatory bacteria: a potential ally against multidrug-resistant
Gram-negative pathogens. _PLoS ONE_ 8, e63397 (2013). Article ADS CAS PubMed PubMed Central Google Scholar * Atterbury, R. J. et al. Effects of orally administered _Bdellovibrio
bacteriovorus_ on the well-being and _Salmonella_ colonization of young chicks. _Appl. Environ. Microbiol._ 77, 5794–5803 (2011). Article ADS CAS PubMed PubMed Central Google Scholar *
Shatzkes, K. et al. Predatory bacteria attenuate _Klebsiella pneumoniae_ burden in rat lungs. _MBio_. 7, e01847-16 (2016). Article PubMed PubMed Central Google Scholar * Willis, A. R.
et al. Injections of predatory bacteria work alongside host immune cells to treat _Shigella_ infection in zebrafish larvae. _Curr. Biol._ 26, 3343–3351 (2016). Article CAS PubMed PubMed
Central Google Scholar * Silhavy, T. J., Kahne, D. & Walker, S. The bacterial cell envelope. _Cold Spring Harb. Perspect. Biol._ 2, a000414 (2010). Article PubMed PubMed Central
Google Scholar * Lerner, T. R. et al. Specialized peptidoglycan hydrolases sculpt the intra-bacterial niche of predatory _Bdellovibrio_ and increase population fitness. _PLoS Pathog._ 8,
e1002524 (2012). Article CAS PubMed PubMed Central Google Scholar * Lambert, C. et al. Interrupting peptidoglycan deacetylation during _Bdellovibrio_ predator-prey interaction prevents
ultimate destruction of prey wall, liberating bacterial-ghosts. _Sci. Rep._ 6, 26010 (2016). Article ADS CAS PubMed PubMed Central Google Scholar * Kuru, E. et al. Fluorescent
D-amino-acids reveal bi-cellular cell wall modifications important for _Bdellovibrio bacteriovorus_ predation. _Nat. Microbiol_. 2, 1648–1657 (2017). Article CAS PubMed PubMed Central
Google Scholar * Harding, C. J. et al. A lysozyme with altered substrate specificity facilitates prey cell exit by the periplasmic predator _Bdellovibrio bacteriovorus_. _Nat. Commun._ 11,
4817 (2020). Article ADS CAS PubMed PubMed Central Google Scholar * Shilo, M. Morphological and physiological aspects of the interaction of _Bdellovibrio_ with host bacteria. _Curr.
Top. Microbiol. Immunol._ 50, 174–204 (1969). Article CAS PubMed Google Scholar * Lambert, C., Fenton, A. K., Hobley, L. & Sockett, R. E. Predatory _Bdellovibrio_ bacteria use
gliding motility to scout for prey on surfaces. _J. Bacteriol._ 193, 3139–3141 (2011). Article CAS PubMed PubMed Central Google Scholar * Matin, A. & Rittenberg, S. C. Kinetics of
deoxyribonucleic acid destruction and synthesis during growth of _Bdellovibrio bacteriovorus_ strain 109D on _Pseudomonas putida_ and _Escherichia coli_. _J. Bacteriol._ 111, 664–673 (1972).
Article CAS PubMed PubMed Central Google Scholar * Hespell, R. B., Miozzari, G. F. & Rittenberg, S. C. Ribonucleic acid destruction and synthesis during intraperiplasmic growth of
_Bdellovibrio bacteriovorus_. _J. Bacteriol._ 123, 481–491 (1975). Article CAS PubMed PubMed Central Google Scholar * Ruby, E. G., McCabe, J. B. & Barke, J. I. Uptake of intact
nucleoside monophosphates by _Bdellovibrio bacteriovorus_ 109J. _J. Bacteriol._ 163, 1087–1094 (1985). Article CAS PubMed PubMed Central Google Scholar * Wyckoff, T. J., Taylor, J. A.
& Salama, N. R. Beyond growth: novel functions for bacterial cell wall hydrolases. _Trends Microbiol._ 20, 540–547 (2012). Article CAS PubMed PubMed Central Google Scholar * Sycuro,
L. K. et al. Flow cytometry-based enrichment for cell shape mutants identifies multiple genes that influence _Helicobacter pylori_ morphology. _Mol. Microbiol._ 90, 869–883 (2013). Article
CAS PubMed PubMed Central Google Scholar * Sycuro, L. K. et al. Peptidoglycan crosslinking relaxation promotes _Helicobacter pylori’s_ helical shape and stomach colonization. _Cell_
141, 822–833 (2010). Article CAS PubMed PubMed Central Google Scholar * Sycuro, L. K. et al. Multiple peptidoglycan modification networks modulate _Helicobacter pylori’_s cell shape,
motility, and colonization potential. _PLoS Pathog._ 8, e1002603 (2012). Article CAS PubMed PubMed Central Google Scholar * Frirdich, E. et al. Peptidoglycan LD-carboxypeptidase Pgp2
influences _Campylobacter jejuni_ helical cell shape and pathogenic properties and provides the substrate for the DL-carboxypeptidase Pgp1. _J. Biol. Chem._ 289, 8007–8018 (2014). Article
CAS PubMed PubMed Central Google Scholar * Frirdich, E. et al. Peptidoglycan-modifying enzyme Pgp1 is required for helical cell shape and pathogenicity traits in _Campylobacter jejuni_.
_PLoS Pathog._ 8, e1002602 (2012). Article CAS PubMed PubMed Central Google Scholar * Min, K. et al. Peptidoglycan reshaping by a noncanonical peptidase for helical cell shape in
_Campylobacter jejuni_. _Nat. Commun._ 11, 458 (2020). Article ADS CAS PubMed PubMed Central Google Scholar * Ausmees, N., Kuhn, J. R. & Jacobs-Wagner, C. The bacterial
cytoskeleton: an intermediate filament-like function in cell shape. _Cell_ 115, 705–713 (2003). Article CAS PubMed Google Scholar * Cabeen, M. T. et al. Bacterial cell curvature through
mechanical control of cell growth. _EMBO J._ 28, 1208–1219 (2009). Article CAS PubMed PubMed Central Google Scholar * Bartlett, T. M. et al. A periplasmic polymer curves _Vibrio
cholerae_ and promotes pathogenesis. _Cell_ 168, 172–185. e115 (2017). Article CAS PubMed PubMed Central Google Scholar * Martin, N. R. et al. CrvA and CrvB form a curvature-inducing
module sufficient to induce cell-shape complexity in Gram-negative bacteria. _Nat. Microbiol_. 6, 910–920 (2021). Article CAS PubMed PubMed Central Google Scholar * Almagro Armenteros,
J. J. et al. SignalP 5.0 improves signal peptide predictions using deep neural networks. _Nat. Biotechnol._ 37, 420–423 (2019). Article CAS PubMed Google Scholar * Kim, H. S. et al. The
cell shape-determining Csd6 protein from _Helicobacter pylori_ constitutes a new family of L,D-carboxypeptidase. _J. Biol. Chem._ 290, 25103–25117 (2015). Article CAS PubMed PubMed
Central Google Scholar * Raghunathan, D. et al. Engulfment, persistence and fate of _Bdellovibrio bacteriovorus_ predators inside human phagocytic cells informs their future therapeutic
potential. _Sci. Rep._ 9, 4293 (2019). Article ADS PubMed PubMed Central Google Scholar * Vollmer, W., Joris, B., Charlier, P. & Foster, S. Bacterial peptidoglycan (murein)
hydrolases. _FEMS Microbiol. Rev._ 32, 259–286 (2008). Article CAS PubMed Google Scholar * Bahadur, R., Chodisetti, P. K. & Reddy, M. Cleavage of Braun’s lipoprotein Lpp from the
bacterial peptidoglycan by a paralog of l,d-transpeptidases, LdtF. _Proc. Natl Acad. Sci. USA_ 118, e201989118 (2021). Article Google Scholar * Winkle, M. et al. DpaA detaches Braun’s
lipoprotein from peptidoglycan. _MBio_. 12, e00836-21 (2021). Article PubMed PubMed Central Google Scholar * Eberhardt, R. Y. et al. Filling out the structural map of the NTF2-like
superfamily. _BMC Bioinformatics_ 14, 327 (2013). Article PubMed PubMed Central Google Scholar * Fenton, A. K., Hobley, L., Butan, C., Subramaniam, S. & Sockett, R. E. A
coiled-coil-repeat protein ‘Ccrp’ in _Bdellovibrio bacteriovorus_ prevents cellular indentation, but is not essential for vibroid cell morphology. _FEMS Microbiol. Lett._ 313, 89–95 (2010).
Article CAS PubMed Google Scholar * Taylor, J. A. et al. Distinct cytoskeletal proteins define zones of enhanced cell wall synthesis in _Helicobacter pylori_. _elife_ 9, e52482 (2020).
Article CAS PubMed PubMed Central Google Scholar * Rittenberg, S. C. Nonidentity of _Bdellovibrio bacteriovorus_ strains 109D and 109J. _J. Bacteriol._ 109, 432–433 (1972). Article CAS
PubMed PubMed Central Google Scholar * Cotter, T. W. & Thomashow, M. F. Identification of a _Bdellovibrio bacteriovorus_ genetic locus, _hit_, associated with the host-independent
phenotype. _J. Bacteriol._ 174, 6018–6024 (1992). Article CAS PubMed PubMed Central Google Scholar * Young, K. D. The selective value of bacterial shape. _Microbiol. Mol. Biol. Rev._
70, 660–703 (2006). Article PubMed PubMed Central Google Scholar * Yang, D. C., Blair, K. M. & Salama, N. R. Staying in shape: the impact of cell shape on bacterial survival in
diverse environments. _Microbiol. Mol. Biol. Rev._ 80, 187–203 (2016). Article CAS PubMed PubMed Central Google Scholar * Wu, F., van Schie, B. G., Keymer, J. E. & Dekker, C.
Symmetry and scale orient Min protein patterns in shaped bacterial sculptures. _Nat. Nanotechnol._ 10, 719–726 (2015). Article ADS CAS PubMed PubMed Central Google Scholar *
Soderstrom, B., Badrutdinov, A., Chan, H. & Skoglund, U. Cell shape-independent FtsZ dynamics in synthetically remodeled bacterial cells. _Nat. Commun._ 9, 4323 (2018). Article ADS
PubMed PubMed Central Google Scholar * Mannik, J. et al. Robustness and accuracy of cell division in _Escherichia coli_ in diverse cell shapes. _Proc. Natl Acad. Sci. USA_ 109, 6957–6962
(2012). Article ADS PubMed PubMed Central Google Scholar * Amir, A., Babaeipour, F., McIntosh, D. B., Nelson, D. R. & Jun, S. Bending forces plastically deform growing bacterial
cell walls. _Proc. Natl Acad. Sci. USA_ 111, 5778–5783 (2014). Article ADS CAS PubMed PubMed Central Google Scholar * Caspi, Y. Deformation of filamentous _Escherichia coli_ cells in a
microfluidic device: a new technique to study cell mechanics. _PLoS ONE_ 9, e83775 (2014). Article ADS PubMed PubMed Central Google Scholar * Mannik, J., Driessen, R., Galajda, P.,
Keymer, J. E. & Dekker, C. Bacterial growth and motility in sub-micron constrictions. _Proc. Natl Acad. Sci. USA_ 106, 14861–14866 (2009). Article ADS PubMed PubMed Central Google
Scholar * Wong, F. et al. Mechanical strain sensing implicated in cell shape recovery in _Escherichia coli_. _Nat. Microbiol_. 2, 17115 (2017). Article CAS PubMed PubMed Central Google
Scholar * Corbett, A. H. & Silver, P. A. The NTF2 gene encodes an essential, highly conserved protein that functions in nuclear transport in vivo. _J. Biol. Chem._ 271, 18477–18484
(1996). Article CAS PubMed Google Scholar * Mitchell, A. L. et al. InterPro in 2019: improving coverage, classification and access to protein sequence annotations. _Nucleic Acids Res._
47, D351–D360 (2019). Article CAS PubMed Google Scholar * Lin, C. S.-H. et al. Peptidoglycan binding by a pocket on the accessory NTF2-domain of Pgp2 directs helical cell shape of
_Campylobacter jejuni_. _J. Biol. Chem_. 296, 100528 (2021). Article CAS PubMed PubMed Central Google Scholar * Lambert, C. & Sockett, R. E. Laboratory maintenance of
_Bdellovibrio_. _Curr. Protoc. Microbiol_. 2, Unit7B.2, 1–14 (2008). * Gibson, D. G. et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. _Nat. Methods_ 6, 343–345
(2009). Article CAS PubMed Google Scholar * Lambert, C. & Sockett, R. E. Nucleases in _Bdellovibrio bacteriovorus_ contribute towards efficient self-biofilm formation and eradication
of preformed prey biofilms. _FEMS Microbiol. Lett._ 340, 109–116 (2013). Article CAS PubMed Google Scholar * Mukherjee, S., Brothers, K. M., Shanks, R. M. Q. & Kadouri, D. E.
Visualizing _Bdellovibrio bacteriovorus_ by using the tdTomato fluorescent protein. _Appl. Environ. Microbiol._ 82, 1653–1661 (2015). Article ADS PubMed Google Scholar * Glauner, B.
Separation and quantification of muropeptides with high-performance liquid chromatography. _Anal. Biochem._ 172, 451–464 (1988). Article CAS PubMed Google Scholar * Peters, K. et al. The
redundancy of peptidoglycan carboxypeptidases ensures robust cell shape maintenance in _Escherichia coli_. _MBio_ 7, e00819-16 (2016). Google Scholar * Bui, N. K. et al. The peptidoglycan
sacculus of _Myxococcus xanthus_ has unusual structural features and is degraded during glycerol-induced myxospore development. _J. Bacteriol._ 191, 494–505 (2009). Article CAS PubMed
Google Scholar * Kabsch, W. XDS. _Acta Crystallogr. D. Biol. Crystallogr._ 66, 125–132 (2010). Article CAS PubMed PubMed Central Google Scholar * Winter, G. xia2: an expert system for
macromolecular crystallography data reduction. _J. Appl. Crystallogr._ 43, 186–190 (2010). Article CAS Google Scholar * Skubak, P. & Pannu, N. S. Automatic protein structure solution
from weak X-ray data. _Nat. Commun._ 4, 2777 (2013). Article ADS PubMed Google Scholar * Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. _Acta Crystallogr.
D. Biol. Crystallogr._ 60, 2126–2132 (2004). Article PubMed Google Scholar * Zwart, P. H. et al. Automated structure solution with the PHENIX suite. _Methods Mol. Biol._ 426, 419–435
(2008). Article CAS PubMed Google Scholar * Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. _Nat. Methods_ 9, 676–682 (2012). Article CAS PubMed
Google Scholar * Ducret, A., Quardokus, E. M. & Brun, Y. V. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. _Nat. Microbiol_. 1, 16077 (2016).
Article CAS PubMed PubMed Central Google Scholar * Lambert, C. et al. Characterizing the flagellar filament and the role of motility in bacterial prey-penetration by _Bdellovibrio
bacteriovorus_. _Mol. Microbiol._ 60, 274–286 (2006). Article CAS PubMed PubMed Central Google Scholar * Lambert, C., Smith, M. C. M. & Sockett, R. E. A novel assay to monitor
predator-prey interactions for _Bdellovibrio bacteriovorus_ 109J reveals a role for methyl-accepting chemotaxis proteins in predation. _Environ. Microbiol._ 5, 127–132 (2003). Article CAS
PubMed Google Scholar * Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. Protein measurement with the Folin phenol reagent. _J. Biol. Chem._ 193, 265–275 (1951). Article
CAS PubMed Google Scholar * Medina, A. A., Shanks, R. M. & Kadouri, D. E. Development of a novel system for isolating genes involved in predator-prey interactions using host
independent derivatives of _Bdellovibrio bacteriovorus_ 109J. _BMC Microbiol._ 8, 33 (2008). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS This work
was funded by a Wellcome Trust PhD studentship (215025/Z/18/Z) to E.J.B., a Becas Chile studentship (72180329) to M.V-D., a BBSRC studentship to A.W., and the UKRI Strategic Priorities Fund
(https://www.ukri.org) EP/T002778/1 to W.V. R.E.S., C.L., and A.L.L. are currently funded by a Wellcome Trust Investigator Award in Science (209437/Z/17/Z). We thank Chloe Hudson for trials
of Bd1075 purification, Daniela Vollmer for purification of PG, Joe Gray for mass spectrometry analysis of muropeptides, Rob Till for general laboratory support, and Block Allocation Group
mx19880 for access to Diamond Synchrotron. Genome sequencing was provided by MicrobesNG (http://www.microbesng.uk), which is supported by the BBSRC (grant number BB/L024209/1). Electron
microscopy was carried out at the Nanoscale and Microscale Research Centre at the University of Nottingham. AUTHOR INFORMATION Author notes * Ian T. Cadby Present address: Bristol Veterinary
School, University of Bristol, Langford, Bristol, BS40 5DU, UK AUTHORS AND AFFILIATIONS * Medical School, School of Life Sciences, University of Nottingham, Queen’s Medical Centre,
Nottingham, NG7 2UH, UK Emma J. Banks, Carey Lambert & R. Elizabeth Sockett * Institute for Microbiology and Infection, School of Biosciences, University of Birmingham, Birmingham, B15
2TT, UK Mauricio Valdivia-Delgado, Amber Wilson, Ian T. Cadby & Andrew L. Lovering * Center for Bacterial Cell Biology, Biosciences Institute, Newcastle University, Newcastle upon Tyne,
NE2 4AX, UK Jacob Biboy & Waldemar Vollmer Authors * Emma J. Banks View author publications You can also search for this author inPubMed Google Scholar * Mauricio Valdivia-Delgado View
author publications You can also search for this author inPubMed Google Scholar * Jacob Biboy View author publications You can also search for this author inPubMed Google Scholar * Amber
Wilson View author publications You can also search for this author inPubMed Google Scholar * Ian T. Cadby View author publications You can also search for this author inPubMed Google
Scholar * Waldemar Vollmer View author publications You can also search for this author inPubMed Google Scholar * Carey Lambert View author publications You can also search for this author
inPubMed Google Scholar * Andrew L. Lovering View author publications You can also search for this author inPubMed Google Scholar * R. Elizabeth Sockett View author publications You can also
search for this author inPubMed Google Scholar CONTRIBUTIONS E.J.B. made all deletion, complementation and fluorescent strains of Bd1075, carried out curvature analysis and conducted
electron, time-lapse, and epifluorescence microscopy experiments with supervision by R.E.S. and C.L. J.B. carried out PG sacculi purifications and HPLC analysis with supervision by W.V.
I.T.C. made the _bd1075_ expression construct for protein purification. A.W. purified the protein with purification optimization and protocol design from I.T.C. M.V-D. crystallized and
solved the structure of Bd1075. All protein work was supervised by A.L.L. E.J.B., R.E.S., and A.L.L. wrote the manuscript and all co-authors read and approved the final manuscript.
CORRESPONDING AUTHORS Correspondence to Andrew L. Lovering or R. Elizabeth Sockett. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER
REVIEW INFORMATION _Nature Communications_ thanks Hesper Rego and the other, anonymous, reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION
SUPPLEMENTARY INFORMATION REPORTING SUMMARY PEER REVIEW FILE DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY MOVIE 1 SUPPLEMENTARY MOVIE 2 SUPPLEMENTARY MOVIE 3 SOURCE DATA
SOURCE DATA RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution
and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if
changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the
material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to
obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS
ARTICLE Banks, E.J., Valdivia-Delgado, M., Biboy, J. _et al._ Asymmetric peptidoglycan editing generates cell curvature in _Bdellovibrio_ predatory bacteria. _Nat Commun_ 13, 1509 (2022).
https://doi.org/10.1038/s41467-022-29007-y Download citation * Received: 24 June 2021 * Accepted: 22 February 2022 * Published: 21 March 2022 * DOI:
https://doi.org/10.1038/s41467-022-29007-y SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative