Play all audios:
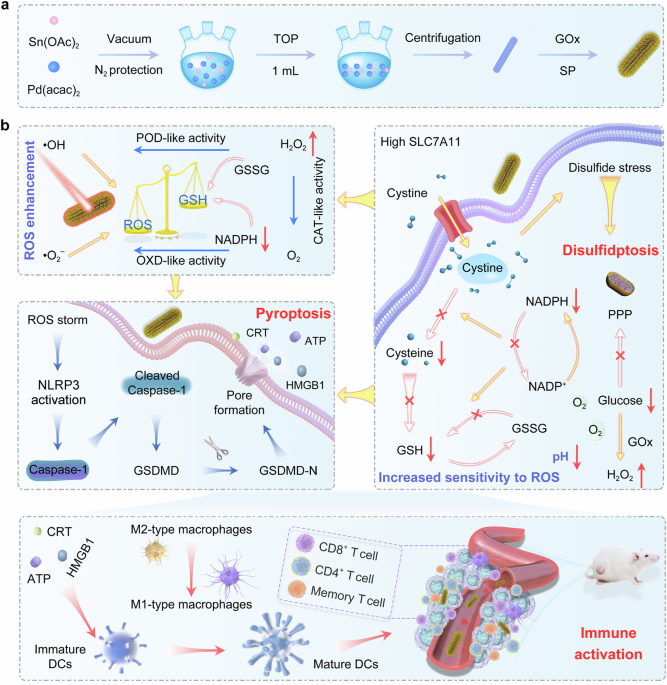
ABSTRACT Pyroptosis, an immunogenic programmed cell death, could efficiently activate tumor immunogenicity and reprogram immunosuppressive microenvironment for boosting cancer immunotherapy.
However, the overexpression of SLC7A11 promotes glutathione biosynthesis for maintaining redox balance and countering pyroptosis. Herein, we develop intermetallics modified with glucose
oxidase (GOx) and soybean phospholipid (SP) as pyroptosis promoters (Pd2Sn@GOx-SP), that not only induce pyroptosis by cascade biocatalysis for remodeling tumor microenvironment and
facilitating tumor cell immunogenicity, but also trigger disulfidptosis mediated by cystine accumulation to further promote tumor pyroptosis in female mice. Experiments and density
functional theory calculations show that Pd2Sn nanorods with an intermediate size exhibit stronger photothermal and enzyme catalytic activity compared with the other three morphologies
investigated. The peroxidase-mimic and oxidase-mimic activities of Pd2Sn cause potent reactive oxygen species (ROS) storms for triggering pyroptosis, which could be self-reinforced by
photothermal effect, hydrogen peroxide supply accompanied by glycometabolism, and oxygen production from catalase-mimic activity of Pd2Sn. Moreover, the increase of NADP+/NADPH ratio induced
by glucose starvation could pose excessive cystine accumulation and inhibit glutathione synthesis, which could cause disulfidptosis and further augment ROS-mediated pyroptosis,
respectively. This two-pronged treatment strategy could represent an alternative therapeutic approach to expand anti-tumor immunotherapy. SIMILAR CONTENT BEING VIEWED BY OTHERS A
HIGH-VALENCE BISMUTH(V) NANOPLATFORM TRIGGERS CANCER CELL DEATH AND ANTI-TUMOR IMMUNE RESPONSES WITH EXOGENOUS EXCITATION-FREE ENDOGENOUS H2O2- AND O2-INDEPENDENT ROS GENERATION Article Open
access 20 January 2025 PROGRAMMED ENHANCEMENT OF ENDOGENOUS IRON-MEDIATED LYSOSOMAL MEMBRANE PERMEABILIZATION FOR TUMOR FERROPTOSIS/PYROPTOSIS DUAL-INDUCTION Article Open access 28 March
2025 PROVOKING TUMOR DISULFIDPTOSIS BY SINGLE-ATOM NANOZYME _VIA_ REGULATING CELLULAR ENERGY SUPPLY AND REDUCING POWER Article Open access 26 May 2025 INTRODUCTION Cancer is one of the
leading causes of death worldwide and exhibits an increasing shift from elderly to middle-aged individuals, in which colorectal cancer is predominant among adults younger than 50 years1,2,3.
Owing to the metabolic reprogramming and genetic mutation, cancer cells usually possess elevated oxidative stress characteristics compared with those of nonmalignant cells4,5,6. To pose the
excessive production of reactive oxygen species (ROS) is detrimental to cancer cells, which has been confirmed as an efficacious cancer therapy. While cancer cells tend to maintain
sufficient glutathione (GSH) levels to neutralize ROS for achieving cell survival and proliferation7. Therefore, blocking the synthesis of GSH is expected to achieve maximum cancer cell
death. Cysteine is a crucial amino acid that affords the rate-limiting precursor for the biosynthesis of GSH8,9. In general, the overexpressed solute carrier family 7 member 11 (SLC7A11) in
most cancer cells could import extracellular cystine (an oxidized cysteine dimer), which could be reduced into cysteine in the cytoplasm with the assistance of glucose-derived nicotinamide
adenine phosphate (NADPH). The cysteine could serve as a precursor for the subsequent synthesis of GSH to achieve antioxidant defense8,10. Thus, the inhibition of NADPH supply could trigger
the accumulation of cystine, thereby leading to the reduction of GSH10. Meanwhile, the overloading of cystine could endow actin cytoskeleton proteins with abundant disulfides and disulfide
bonds, resulting in disulfidptosis caused by disulfide stress6,11. Besides the suppression of GSH, promoting massive ROS production also plays a critical role in intensive tumor treatments.
The slightly acid and overexpressed hydrogen peroxide (H2O2) of tumor microenvironment (TME) create excellent conditions for the production of ROS catalyzed by nanocatalysts12.
Intermetallics composed of two or more metallic elements display expanded and superior catalytic activity owing to the tuned electronic states and the synergistic effect between the two
metals13,14. Significantly, the positions of all atoms in intermetallics are assigned with specific sites, leading to the defined stoichiometry and crystal structure15. Compared with common
alloys with occupy random sites, the regular structure endows the intermetallics with homogeneity of the active sites, which could ensure the abundant generation of ROS16. In addition to the
enhancement of catalytic activity, the stability of intermetallics could also be improved, which makes them perform better in various physiological environments. This enhancement could be
attributed to the mixed bonding, which leads to a more negative free energy of formation than random alloys17. Besides the ordered structure, the shape of intermetallics has an influence on
the crystal plane and active sites, which could also determine the catalyst performance18. Thus, developing well-controlled intermetallics with tuned size and shape is attractive in
improving the catalytic capacity and optimizing the formation of ROS for effective cancer therapeutics. Furthermore, previous research has validated the decisive role of ROS in inducing
immunogenic programmed cell death, which could efficiently arouse acute inflammatory response and trigger robust anti-tumor immune activity19,20,21. Characterized by the activation of
Caspase-1 and cleavage of gasdermin D (GSDMD), pyroptosis is an inflammatory cell death. The N-terminal domain of GSDMD (N-GSDMD) could perforate the plasma membrane and induce cell membrane
rupture, releasing the bountiful pro-inflammatory cytokine and tumor antigens, which could provoke precision cancer immunotherapy22,23,24. Delightedly, a low level of pyroptosis (<15%)
in tumor cells could achieve an efficacious anti-tumor immunity25. Consequently, the ROS-induced pyroptosis could not only kill cancer cells by cell membrane perforation, but also facilitate
the initiation and infiltration of T cells owing to the release of cytoplasmic contents, which could remodel the tumor immunosuppressive microenvironment and transform cold tumor
(immune-indolent noninflamed tumor phenotype) into hot tumors (inflammatory phenotype)24,26. Therefore, exploring the effective strategies to amplify oxidative stress-triggered pyroptosis
while synchronously magnifying immunity responses in the TME offers a distinctive direction for cancer therapy. In this work, we report the intermetallics modified with glucose oxidase (GOx)
and soybean phospholipid (SP) as nano-inducers (Pd2Sn@GOx-SP) to elicit potent anti-tumor immune responses by pyroptosis and disulfidptosis (Fig. 1). To begin with, the Pd2Sn intermetallics
with different morphologies are developed and their performance are contrasted both in experiments and density functional theory (DFT) calculations. Among them, the Pd2Sn nanorods (NRs)
with the largest specific surface area exhibit the optimal photothermal and enzyme catalytic activities, including peroxidase (POD)-mimic and catalase (CAT)-mimic catalytic activity. In
addition, the modification of GOx on the Pd2Sn intermetallics could serve as a triple role: (1) degrade glucose to afford H2O2 for the cascade catalysis reaction, triggering a strong ROS
storm; (2) reduce GSH synthesis to defeat the antioxidant defense mechanism of cells and aggravate oxidative stress; (3) inhibit NADPH supply by regulating the glycometabolism to induce the
accumulation of cystine, resulting in disulfidptosis. Moreover, the CAT-mimic catalytic activity of Pd2Sn intermetallics could effectively restore the O2 levels and further enhance the
breakdown of glucose, thereby achieving a self-promoted process. This potent and persistent cellular oxidative pressure could effectively activate and amplify ROS-mediated pyroptosis. In
summary, the integration of Pd2Sn intermetallics and GOx could arouse self-reinforced ROS storm and glucose consumption, which could cause Caspase-1-dependent pyroptosis and cystine-mediated
disulfidptosis, leading to the reprogramming of immune microenvironment. Meanwhile, the change of immunosuppressive microenvironment, including the ameliorative infiltration of T cells
(CD4+ and CD8+) and M1-like phenotype repolarization of macrophages, could achieve efficacious immunotherapy by activating immune response, counteracting tumor recurrence and metastasis.
RESULTS CONSTRUCTION AND CHARACTERIZATION OF PD2SN@GOX-SP NANOCOMPOSITES The intermetallic compounds of Pd2Sn nanoparticles with different morphologies were synthesized through one step by
the co-reduction of palladium(II) acetylacetonate [Pd(acac)2] and tin(II) acetate [Sn(OAc)2], as shown in Fig. 1a. Methylamine hydrochloride (MAHC) played an important role in controlling
the morphology of Pd2Sn intermetallics, including nanodots (NDs, without MAHC) and NRs in different sizes (with different amounts of MAHC). The representative transmission electron
microscopy (TEM) micrographs and elemental mapping of the Pd2Sn intermetallics were shown in Fig. 2a, b and Supplementary Fig. 2a–c, respectively. The Pd2Sn intermetallics exhibited the
uniform controllable morphology of NRs and NDs, as well as the homogeneous distribution of corresponding elements, confirming the successful construction of intermetallics with different
morphologies. High-resolution TEM (HRTEM) of the Pd2Sn intermetallics revealed the interfacial lattice spacings of 0.195 nm (NRs) and 0.236 nm (NDs), which could be identified as (3 2 0) and
(0 2 1) crystal planes of the orthorhombic phase, respectively (Fig. 2c, d and Supplementary Fig. 2d, e). In addition, the coexistence of Pd and Sn elements (with a ratio of 2:1) could also
be observed from the energy-dispersive spectroscopy (EDS) spectrum (Fig. 2e), suggesting the successful synthesis of Pd2Sn intermetallics. Powder X-ray diffraction (PXRD) patterns of Pd2Sn
intermetallics were presented in Fig. 2f and Supplementary Fig. 2, the diffraction peaks showed good crystallinity with the lattice parameter of _a_ = 8.11 Å, _b_ = 5.662 Å, _c_ = 4.234 Å,
which matched well with the pure orthorhombic Pd2Sn phase (PDF 00-026-1297). The Pd2Sn with two inequivalent Pd sites is cotunnite structured and crystallizes in the orthorhombic Pnma space
group (Fig. 2g). Additionally, X-ray photoelectron spectroscopy (XPS) was performed to analyze the composition and valence state information of Pd2Sn intermetallics (NDs and NRs). The
as-prepared Pd2Sn intermetallics were mainly composed of Pd, Sn, and O elements (Fig. 2h). The Pd 3_d_ region of Pd2Sn intermetallics in the high-resolution XPS spectrum could be assigned to
the spin-splitting orbits of 3_d_3/2 and 3_d_5/2, and the binding energies at 341.0 and 335.9 eV were attributed to Pd2+, while the peaks at 340.1 and 335.0 eV were assigned to Pd0 (Fig.
2i). This result confirmed the coexistence of Pd0 (77%) and Pd2+ (23%) in Pd2Sn NRs and Pd0 (74%) and Pd2+ (26%) in Pd2Sn NDs. Meanwhile, the high-resolution XPS spectrum for Sn 3_d_ of
Pd2Sn intermetallics exhibited a distinct decomposition, verifying the existence of tin with two distinct valence states (Fig. 2j). The characteristic peaks at 486.6 eV (3_d_5/2) and 497.4
eV (3_d_3/2) were attributed to Sn4+, while the peaks at 486.0 eV (3_d_5/2) and 496.7 eV (3_d_3/2) were associated with Sn2+, and the component at 484.5 eV (3_d_5/2) and 494.8 eV (3_d_3/2)
corresponded to metallic Sn. Furthermore, we also investigated the change of elements in the valence state after prolonged oxidation (Supplementary Fig. 3a, b). The majority of the surface
Sn was in the oxidized state and the surface Pd was mainly in the metallic state, indicating the surface Sn atoms are more easily oxidized than Pd atoms. These results validated that the
Pd2Sn intermetallics could generate more ROS through the redox reaction27. The corresponding elemental mapping, EDS spectrum, and Fourier transform infrared spectrum (FT-IR) showed that the
surface of Pd2Sn intermetallics interacted with Tri-n-octylphosphine (TOP) due to the coordination potence of phosphine ligands (Supplementary Figs. 4 and 5a). To endow the Pd2Sn
intermetallics with high biocompatibility, the SP was further functionalized on the surface of Pd2Sn intermetallics to obtain Pd2Sn-SP nanocomposites28. Significantly, the surface of Pd2Sn
intermetallics was further modified with GOx, which could facilitate the cascade catalytic reaction to generate more ROS by the enhancement of H2O2 catalyzed by GOx. The successful
functionalization of SP and modification of GOx were confirmed by the FT-IR spectrum (Supplementary Fig. 5b). Thermogravimetric analysis (TGA) confirmed that the proportion of SP on Pd2Sn-SP
nanocomposites was ~20.81% (Supplementary Fig. 6). In addition, the zeta potentials showed obvious changes from 0.13 to −29.93 mV after the functionalization of SP, and the zeta potential
of Pd2Sn@GOx-SP was further changed to −23.0 mV (Supplementary Fig. 7a). Furthermore, the hydrodynamic size of Pd2Sn@GOx-SP nanocomposites in phosphate buffer saline (PBS) was ~90 nm
according to the analysis of dynamic light scattering, and the average size of Pd2Sn@GOx-SP at various solvents was also evaluated (Supplementary Fig. 7b). The Pd2Sn@GOx-SP exhibited a
uniform hydrated particle size over 7 days, demonstrating the ideal stability of Pd2Sn@GOx-SP in physiological conditions, which is a prerequisite for better subsequent application.
PHOTOTHERMAL AND MULTI-ENZYMATIC CATALYTIC ACTIVITY EVALUATION As a noble metal, palladium exhibits the peculiar localized surface plasmon resonance (LSPR) effect and various enzyme
catalytic activity, which has made a profound study (Fig. 3a)29,30,31,32. To begin with, the optical properties of Pd2Sn-SP nanocomposites (NDs and NRs) were explored by using UV–vis–NIR
absorbance spectrum (Fig. 3b and Supplementary Fig. 8), the adsorption intensity of Pd2Sn-SP NDs and NRs increased linearly with an elevation of concentration. Especially, the Pd2Sn-SP NRs
exhibited stronger absorption (5.33 L g−1 cm−1) than Pd2Sn-SP NDs with the extinction coefficient was 2.30 L g−1 cm−1. Subsequently, the Pd2Sn-SP NRs were chosen for evaluating the
photothermal performance. A notable laser power density-dependent photothermal effect was observed in Supplementary Fig. 9, and the temperature of Pd2Sn-SP NRs irradiated with laser (0.8 W
cm−2) can considerably increase by 26 °C, which is sufficient for killing cancer cells. It is noteworthy that the temperature change of Pd2Sn-SP NRs also showed concentration-dependent (Fig.
3c), while only a slight increase of pure water with laser excitation (0.8 W cm−2), which was also verified from the infrared thermal images (Supplementary Fig. 10), indicated the excellent
efficiency of Pd2Sn-SP NRs for photothermal conversion. Besides photothermal conversion capability, photothermal stability is also critical for the photothermal conversion process, which
was further assessed by four on/off laser power cycles. No notable changes in temperature even after four cycles of heating and natural cooling, confirming high photothermal stability (Fig.
3d). Then, the photothermal conversion efficiency (_η_) of Pd2Sn-SP NRs was calculated to be 41.2% on the basis of cooling phase (Fig. 3e). The high light absorption and excellent
photothermal conversion ability of Pd2Sn-SP NRs make them effective contrast agents for photoacoustic imaging (PAI) due to the LSPR effect33,34. Consequently, the in vitro PAI ability of
Pd2Sn-SP NRs was explored by detecting the PA signals of Pd2Sn-SP NRs with different concentrations. As displayed in Supplementary Fig. 11, the PA signals showed a positive correlation with
the concentration of Pd2Sn-SP NRs, which demonstrated the ideal PAI ability for biomedical and clinical applications. Taking the excellent catalytic activity for effective anti-cancer
treatment into consideration, the multienzyme-mimic activities as well as the cascade catalytic reaction of Pd2Sn-SP nanocomposites were explored32,35. To begin with, the generation of 1O2
and ·OH was quantitatively evaluated by the electron spin resonance (ESR) measurement with 2,2,6,6-tetramethylpiperidine (TEMP) and 5,5-dimethyl-1-pyrroline _N_-oxide (DMPO) as trapping
agents (Fig. 3f). An obvious signal peak of 1O2 could be seen when the Pd2Sn-SP nanocomposites were irradiated with a laser, and the stronger signals occurred after the addition of H2O2
(Supplementary Fig. 12a). The distinct quadruple resonance peaks of ·OH could be observed in Fig. 3g, displaying a signal intensity ratio of 1:2:2:1, which was further enhanced upon laser
irradiation, indicating the laser-induced hyperthermia-enhanced production of ·OH. Moreover, 9,10-diphenylanthracene (DPA) was further used to confirm the generation of 1O2, the absorption
intensity of DPA weakened with time after being exposed to the laser, illustrating the generation of 1O2 by Pd2Sn-SP nanocomposites (Supplementary Fig. 12b). Simultaneously, the POD-mimic
activity of Pd2Sn-SP NRs was also validated by the color reaction of _o_-phenylenediamine (OPD) and 3,3′,5,5′-tetramethyl-benzidine (TMB), which could be oxidized by ·OH to generate orange
(oxOPD) or blue color (oxTMB) with the characteristic peak at ~420 or ~652 nm, respectively. The Pd2Sn-SP NRs induced remarkable characteristic absorption of oxOPD or oxTMB after adding
H2O2, while the control or laser alone group showed no changes (Fig. 3h, i). Moreover, the absorption intensity was enhanced when the Pd2Sn-SP NRs irradiated with a laser, which was similar
to the results of ESR measurement. Furthermore, the difference in catalytic activity of Pd2Sn-SP nanocomposites with diverse morphology (NDs, NRs, SNRs, and LNRs) was investigated by the
steady-state kinetic analyses. Four types of Pd2Sn-SP nanocomposites were mixed with H2O2 at various concentrations (1, 2, 4, 8, and 16 mM), and the absorbance change of oxTMB was recorded
with time (Supplementary Fig. 13), all of which could align well with the typical Michaelis–Menten kinetics (Fig. 3j). Based on the Lineweaver–Burk plots (Fig. 3k), the maximum reaction
velocity (_V_max) and Michaelis–Menten constants (_K_m) were acquired and summarized in Fig. 3l. Generally, a higher _V_max and lower _K_m lead to better catalytic performance. Compared with
the Pd2Sn-SP NDs, the Pd2Sn-SP NRs exhibited higher catalytic efficiency. Moreover, the catalytic efficiency was also relevant to the length of Pd2Sn-SP NRs, neither too long nor too short
is beneficial for the POD-mimic activity. Additionally, Pd2Sn-SP nanocomposites with diverse morphology were employed to further evaluate the catalytic activity by altering the
concentrations of Pd2Sn-SP nanocomposites and detecting the absorption intensity of oxTMB (Supplementary Fig. 14a–d). The specific activity values of Pd2Sn-SP nanocomposites were calculated
(Supplementary Fig. 14e–h) and summarized in Supplementary Fig. 14i. Similarly, the Pd2Sn-SP NRs showed the supreme POD-mimic catalytic activity than the others. The effective generation of
·OH in the presence of Pd2Sn-SP NRs was also confirmed by performing a methylene blue (MB) experiment. As displayed in Supplementary Fig. 15, the absorbance of MB decreased with the
prolonged reaction time, demonstrating the excellent generation ability of ·OH. In addition to ·OH, the ·O2− generation based on the oxidase (OXD)-mimic catalytic activity of Pd2Sn-SP NRs
was also validated using TMB (Fig. 3n). The absorbance intensity at 652 nm increased with a prolonged time, showing the obvious OXD-mimic activity of Pd2Sn-SP NRs. Considering the hypoxic
TME restricts the OXD-mimic activity, the CAT-mimic catalytic activity of Pd2Sn-SP NRs by catalyzing H2O2 into O2 was detected. The Pd2Sn-SP nanocomposites with diverse morphology were added
with various concentrations of H2O2 and the oxygen content was recorded for comparison. As shown in Supplementary Fig. 16, all the Pd2Sn-SP nanocomposites could produce more O2 with the
increase of H2O2 concentration, and Pd2Sn-SP NRs displayed the strongest CAT-mimic catalytic activity, which was consistent with the results of POD-mimic catalytic activity. In addition, the
generation of O2 also increased with the concentration of Pd2Sn-SP NRs (Fig. 3o), and the addition of laser further promoted the CAT-mimic catalytic activity (Fig. 3p). Although the TME is
featured by the overexpression of H2O2, the endogenous H2O2 content is unable to meet the subsequent biocatalytic reactions36. Thus, the Pd2Sn-SP NRs were modified with GOx to supply H2O2 by
catalyzing glucose decomposition. The catalytic ability of Pd2Sn@GOx-SP for accelerating glucose consumption was verified, the glucose consumption increased with the prolonged reaction
time, while the negligible change was observed in the control group (Fig. 3q). Meanwhile, the degradation of glucose is accompanied by the production of gluconic acid, leading to the
reduction of pH value (Fig. 3r). Therefore, the consumption of endogenous glucose could not only provide H2O2 for enzyme-catalyzed reaction, but also reduce pH value to enhance the enzyme
catalytic efficiency. Furthermore, the catalytic activity of GOx was evaluated in PBS with different pH values to investigate whether the GOx would malfunction in a more acidic environment.
The results showed that there was no obvious change in the catalytic performance of GOx in a more acidic environment, demonstrating that the GOx was able to effectively degrade glucose and
afford H2O2 for the cascade catalytic reaction (Supplementary Fig. 17). Moreover, the O2 generated by CAT-mimic catalytic activity could also improve the OXD-mimic activity as well as the
glucose consumption by GOx, achieving mutual promotion and common improvement (Fig. 3m). We also evaluated the influence of pH value on the etching and leaking of Sn and Pd ions by using
inductively coupled plasma-optical emission spectrometry (ICP-OES). As shown in Supplementary Fig. 18, the Sn and Pd ions could be released from Pd2Sn@GOx-SP, while the amount of ion release
was negligible even during 3 days of incubation, which was only 2.73 and 1.10 μg mL−1, respectively. The ability to reshape the TME and promote the extensive generation of ROS endows
Pd2Sn@GOx-SP with excellent potentiality for tumor therapy. DFT CALCULATIONS AND ENZYMATIC CATALYSIS MECHANISM DFT calculations were further performed to unveil the catalytic mechanism and
the crystal-facet-dependent multienzyme-mimicking activities of Pd2Sn. First, the bulk structure of Pd2Sn was optimized and displayed in Fig. 4a. According to the calculated density of
states (DOS) in Fig. 4b, it can be identified that the predominant contribution of the DOS near the Fermi level originated from Pd-5_d_ orbitals, indicating that electrons near the Fermi
level are mainly provided by Pd atoms and of great importance for the catalytic activity of Pd2Sn. The moderate atomic radius and _d_ orbitals with abundant electrons of the Pd made Pd2Sn
easy to interact with the reactant molecules and promote the electron transfer process. Thus, the excellent catalytic activities of Pd2Sn could be achieved by activating the reactants and
subsequently forming suitable intermediates with lower energy barriers. Based on the facet discrepancy of Pd2Sn NDs and NRs, three surface models with exposed (001), (010), and (011) crystal
facets were built to clarify the enzyme catalytic mechanisms (Fig. 4c). Before evaluating the catalytic activity of different Pd2Sn facets, microscopic electron structures and Bader charge
analysis were conducted to observe the charge distribution. As shown in Fig. 4d, the surface electrons were transferred from Sn to Pd atoms, which altered the redistribution of electrons and
led to the negative valence state of Pd. It can be observed clearly that the charges of the Pd sites on (001), (101), and (011) surfaces were calculated to be −0.297, −0.418/−0.484, and
−0.261/−0.339/−0.348 e, respectively. The significant change of surface electron density could have an impact on the catalytic performance of Pd2Sn catalysts, as discussed in detail later.
Furthermore, to evaluate the electron transfer behavior between the catalyst surface and the intermediates, the work functions of different facets were also calculated37. In response to the
surface charge redistribution, the data in Fig. 4e–g identified that the work functions for (001), (010), and (011) facets are 6.902, 6.341, and 7.241, indicating the electron transfer from
the catalyst surface to the intermediates followed by the sequence of (010) > (001) > (011). In addition, the adsorption process of H2O and H2O2 on the Pd and Sn sites of different
Pd2Sn facets was analyzed and summarized in Supplementary Fig. 19a. The adsorption energy of H2O2 on Pd and Sn sites of three different crystal facets were much lower than that of H2O, which
was beneficial for its preferential adsorption thus the subsequent catalysis (Supplementary Fig. 19b). Significantly, the corresponding adsorption energies of H2O2 on Pd site in (001),
(010), (011) facets were calculated to be −0.28, −0.17, and −0.13 eV, respectively. The lower adsorption energy of H2O2 on the Pd site with respect to the Sn site in the (001) facet ensured
the energetically favorable binding of H2O2 on the catalyst surface. Furthermore, to reveal the relevant catalytic mechanism, three optimized surfaces of Pd2Sn were adopted to
comprehensively investigate the H2O2 catalytic process. The critical intermediates formed in the successive reaction steps of CAT-mimic and POD-mimic activities were displayed in Fig. 4h and
Supplementary Fig. 19c. All the H2O2 catalytic reaction intermediates were optimized for the (001), (010), and (011) crystal facets. Regarding the POD-mimic activity, the calculated energy
profiles are illustrated in Fig. 4i and Supplementary Fig. 19d. The first step involves the capture and surface adsorption of H2O2 on the Pd2Sn surface. Then, the H2O2 was decomposed into
two OH*, which was thermodynamically favorable in the three crystal facets. In the third step, the adsorbed OH* was released and formed ·OH, which required to overcome an energy barrier and
thus became the rate-determining step. Pd sites on the (001) facet possessed the lowest free energy (2.59 eV per OH*) for the desorption of bridged OH* intermediate compared with (010) and
(011) crystal facets (2.73 and 2.63 eV per OH*, respectively), demonstrating its better POD-mimic activity. Simultaneously, the energy profiles for the CAT-mimic activity were shown in Fig.
4j and Supplementary Fig. 19e. Five successive catalytic processes on the (001), (010), and (011) crystal facets have been calculated. The adsorption, activation, and dissociation of H2O2
were all exothermic in all crystal facets, indicating the feasibility of the reaction process. Conversely, the deprotonation of OH* to form O* and the subsequent formation of bridge
structure O2* were apparent endothermic processes. For the Pd site on Pd2Sn (001) surface, the energies required to form O* and O2* intermediates are calculated to be 1.06 eV (deprotonation
of one H) and 1.69 eV, respectively. In contrast, relevant values for Pd2Sn (010)/(011) surfaces are 1.22/1.41 and 0.88/1.89 eV. Moreover, the deprotonation of OH* on the Pd2Sn (001) surface
is much easier than that on the Pd2Sn (010) surface, while the energy required to form O2* on the Pd2Sn (010) surface is the lowest. Thus, the formation of O2* intermediates became the
rate-determining step in the catalytic processes of CAT-mimic activity. It should be noted that the deprotonation of OH* to produce hydrogen products (H2) required an electron transfer
between catalyst surface and intermediates. Therefore, the highest work function of (011) facet would result in the highest interfacial charge transfer resistance during this fundamental
step, even the (011) facet exhibited a much lower energy barrier. In addition, as all the energy barriers needed to overcome for Pd sites on (001) and (010) surface are significantly lower
than the one (1.89 eV) to form O2* on (011) surface, demonstrating the CAT-mimic activity of the Pd2Sn (011) surface is the worst. According to Supplementary Fig. 19f, it can be further
confirmed that the energy barrier for the formation of O2* at Pd sites is well correlated with the work function of different facets, implying that the charge redistribution caused by the
change of the surface orientation will alter the electronic structure of the catalyst surface, and thus changing the reaction mechanism. In addition, the results in Supplementary Fig. 19g
also indicated that the Sn sites in the three facets do not exhibit obvious superiority over the formation of O* and O2* intermediates, but they are favorable for the desorption of O2 with
the energies ranging from −0.19 to 0.10 eV. According to the analysis on the adsorption, reaction, and desorption capabilities of different sites on different facets, it can be concluded
that Pd and Sn sites play different but synergistic roles in determining the overall activity during whole CAT-mimic reaction. In brief, the POD-mimic activity is easier to occur in (001)
crystal facet than in (011) crystal facet. The (001) and (010) crystal facets play different roles in CAT-mimic activity, in which (001) facet facilitates the deprotonation of OH* and (010)
facet facilitates the formation of O2*. Both (001) and (010) crystal facets exhibit better CAT-mimic activity than (011) crystal facets. Moreover, Sn sites facilitate the desorption of O2,
collaboratively promoting the CAT-mimic activity. Overall, the CAT-mimic and POD-mimic activities of Pd2Sn were theoretically verified by the exploration of the reaction pathways and energy
profiles, further demonstrating that the Pd2Sn NRs with exposed (001) facet possessed higher catalytic activity than Pd2Sn NDs with exposed (011) facet. IN VITRO SYNERGISTIC PYROPTOSIS AND
DISULFIDPTOSIS These experiments have confirmed that the Pd2Sn@GOx-SP possessed the property for altering the TME and generating abundant ROS. We further evaluated the in vitro anti-tumor
therapeutic effect of Pd2Sn@GOx-SP (Fig. 5a). To begin with, we measured the cytotoxicity of Pd2Sn@GOx-SP on CT26 cells by methylthiazolyldiphenyl-tetrazolium bromide (MTT) assay. Various
concentrations of Pd2Sn-SP and Pd2Sn@GOx-SP were co-incubated with CT26 cells in the dark to detect the dark-toxicity, and the photo-toxicity of Pd2Sn@GOx-SP was detected by laser
irradiation followed by the incubation. As can be seen in Fig. 5b, both Pd2Sn-SP and Pd2Sn@GOx-SP were cytotoxic to CT26 cells compared with drug-free incubation. Significantly, the
Pd2Sn@GOx-SP exhibited remarkable photo-toxicity on tumor cells under laser irradiation. Moreover, the cytotoxicity of Pd2Sn@GOx-SP nanocomposites on normal cells and multiple cancer cells
was further performed to evaluate the tumor growth inhibitory effect. Our findings revealed that the nanocomposites showed varying degrees of inhibitory effects on cancer cells
(Supplementary Fig. 20a). While for normal cells (L929 and 3T3 cells), both Pd2Sn-SP and Pd2Sn@GOx-SP were almost non-cytotoxic after co-incubation of 12 and 24 h, showing good
biocompatibility even at higher concentrations (Supplementary Fig. 20a–d). Given the existence of TOP on the surface of Pd2Sn, we further co-cultured the L929 cells with TOP at different
concentrations to evaluate the biotoxicity of P originated from TOP. The result showed no significant toxic side effects on L929 cells, demonstrating the negligible biotoxicity of P in TOP
(Supplementary Fig. 20e). Meanwhile, the influence of Sn and Pd ions leaking on L929 cells was also investigated through MTT assay by co-culturing the released Sn and Pd ions with L929
cells. The results showed that there was no obvious biotoxicity on L929 cells (Supplementary Fig. 20f), indicating the high biosafety of Pd2Sn@GOx-SP. Thus, the nanocomposites exhibited
negligible cytotoxicity on normal cells without laser irradiation, demonstrating high selective toxicity. To explore the cellular internalization performance of Pd2Sn@GOx-SP, the cancer
cells incubated with fluorescein isothiocyanate (FITC) modified Pd2Sn@GOx-SP were analyzed by confocal laser scanning microscopy (CLSM) and flow cytometry, respectively (Fig. 5c, d and
Supplementary Fig. 21a, b). The cells treated with FITC-labeled Pd2Sn@GOx-SP showed higher fluorescence over a prolonged incubation time, exhibiting the excellent cellular uptake efficiency
of Pd2Sn@GOx-SP. Research has shown that endocytosis is the main internalization process of nanocomposites, and the uptake rate of rod-like nanocomposites was higher than that of spherical
nanocomposites38,39,40. The endosomes/lysosomes usually play an indispensable role in transporting and releasing nanocomposites during subsequent intracellular processes41. Consequently, the
endosomal escape process of Pd2Sn@GOx-SP was evaluated by staining lysosomes with Lyso-Tracker. The results revealed that the FITC-labeled Pd2Sn@GOx-SP was colocalized with lysosomes within
1 h after internalization (Pearson’s correlation coefficient of 0.79). With the extension of incubation time, the Pd2Sn@GOx-SP gradually escaped from the lysosomes, as evidenced by the
Pearson’s correlation coefficient was decreased to 0.52 and 0.39 after 2 and 4 h of internalization, respectively (Fig. 5c), indicating that the Pd2Sn@GOx-SP could effectively escape from
endolysosomes. Considering stereoscopic structure of the tumor, we further investigated the penetration performance of Pd2Sn@GOx-SP on CT26 multicellular spheroids to simulate
three-dimensional tumors. Similarly, the fluorescence intensity of multicellular spheroids incubated with FITC-labeled Pd2Sn@GOx-SP enhanced with the time prolongs (Supplementary Fig. 21c),
indicating the ideal permeability for the multicellular spheroids. Thus, the ideal penetration ability could promote the accumulation of Pd2Sn@GOx-SP in solid tumors, which could gain more
access to tumor cells and improve the therapeutic efficacy. Subsequently, the change of intracellular substances and their effect on cell death was further evaluated on CT26 cells. The
oxygen content in tumor cells plays a significant role in tumor therapy42. On the one hand, the generated O2 could accelerate the consumption of glucose in the presence of GOx to trigger a
series of cellular damage. On the other hand, the abundant O2 could be converted into 1O2 and ·O2− catalyzed by Pd2Sn@GOx-SP, which enhances the oxidative damage of tumor cells.
[Ru(dpp)3]2+Cl2 as an oxygen detection probe was used to evaluate the oxygen production of Pd2Sn@GOx-SP in CT26 cells. As displayed in Fig. 5e, different from the control and laser groups,
the red fluorescence intensity of the cells decreased after incubation with Pd2Sn-SP, suggesting that the continuous production of O2 based on the CAT-mimic catalytic activity of the
Pd2Sn-SP. It was also comparatively found that the cells in the Pd2Sn@GOx-SP group exhibited a brighter red fluorescence than that of Pd2Sn-SP, which could be attributed to the O2
consumption in the decomposition of glucose by GOx. Whilst, a large amount of H2O2 generation is accompanied by the process of glucose decomposition, which could further augment the content
of ROS. It has been confirmed that the abnormal increase of intracellular ROS levels could induce damage to crucial cellular biomolecules, leading to cell death (including pyroptosis)
through the Caspase-1/GSDMD pathway43,44. Various strategies to generate ROS have been confirmed in vitro and showed excellent anti-tumor therapeutic potential45. Thus, the intracellular ROS
was determined by CLSM and flow cytometry using a 2′,7′-dichlorofluorescein diacetate (DCFH-DA) probe (Fig. 5f and Supplementary Fig. 21d, e). The results showed the fluorescence intensity
of Pd2Sn@GOx-SP group was remarkable, demonstrating the exceptional ability of ROS generation. Especially, the highest fluorescence intensity was observed in the Pd2Sn@GOx-SP + laser group
due to the enhancement of ROS generation by the hyperthermia effect. High levels of ROS in tumor cells could achieve irreversible damage to DNA to induce pyroptosis46. Considering the
abnormal elevated ROS levels could lead to mitochondrial dysfunction, the change of mitochondrial membrane potential after various treatments was evaluated by performing JC-1 staining. As
shown in Supplementary Fig. 21f, JC-1 forms aggregates within the matrix of mitochondria under normal polarization conditions, exhibiting vibrant red fluorescence. On the contrary, JC-1
could only exist as a monomer in depolarized mitochondria, showing green fluorescence46. As depicted in Fig. 5g, intense red fluorescence was shown in the cells with the treatment of control
and only laser, indicating normal mitochondria. In particular, the green fluorescence intensity of cells treated with Pd2Sn@GOx-SP increased significantly, and the highest green–red
fluorescence proportion was exhibited in the Pd2Sn@GOx-SP + laser group, demonstrating severe mitochondrial damage. Meanwhile, highly toxic ROS could analogously destroy the lysosomes47,
which are also crucial subcellular organelle. The integrity of lysosomes was analyzed by acridine orange staining assay, in which red fluorescence could change into green fluorescence when
the lysosome ruptured. The results showed that the lysosomal membrane integrity was completely ruptured after the treatment of Pd2Sn@GOx-SP + laser, and a certain rupture could also be
observed in the Pd2Sn-SP and Pd2Sn@GOx-SP groups (Supplementary Fig. 22a). The destruction of Pd2Sn@GOx-SP on CT26 cells was also confirmed by observing the morphological changes of actin
filaments (F-actin). The cytoskeletal disruption could be seen in the cells after being treated with Pd2Sn@GOx-SP, while the F-actin of the cells in the group of control or only laser were
highly elongated and well-organized (Supplementary Fig. 22b), demonstrating the cell injury induced by Pd2Sn@GOx-SP. To gain further insights into the living and dead cells, Calcein-AM/PI
was used to stain the cells in various groups, and the fluorescence intensity was examined (Supplementary Fig. 22c). The dead cells in the control or only laser group were negligible, while
dead cells predominated in the group of Pd2Sn@GOx-SP + laser, indicating the strong killing efficiency of Pd2Sn@GOx-SP on tumor cells. The living and dead cells proportion in different
treatment groups was also obtained by flow cytometric assay. The dead cells increased after treatment with Pd2Sn-SP or Pd2Sn@GOx-SP, and the cell mortality rate reached its maximum after
incubation with Pd2Sn@GOx-SP + laser (Supplementary Fig. 22d), suggesting the ideal treatment effect of Pd2Sn@GOx-SP on cancer cells. Besides that, we also investigated the therapeutic
performance of Pd2Sn@GOx-SP on CT26 multicellular spheroids in vitro. Consistent with the above results, almost no cell death was observed in control or only laser groups, while a certain
amount of dead cells existed in Pd2Sn@GOx-SP group, and a significant increase of dead cells was found in Pd2Sn@GOx-SP + laser group (Fig. 5h). Different depths of cell death could be
observed, confirming the effective ability for tumor inhibition. After confirming the excellent capability for killing tumor cells, we further explored the specific mechanism of
Pd2Sn@GOx-SP-induced cell death in detail. Due to the massive production of ROS with various types, we speculated that Pd2Sn@GOx-SP could induce pyroptosis46,48,49. As an inflammatory
programmed cell death, pyroptosis could activate the anti-tumor immune responses by releasing various inflammatory factors, which could serve as an important means to improve immune
deficiencies25. To verify the immune activation effect induced by Pd2Sn@GOx-SP, the typical biomarkers of immunogenic cell death (ICD), high mobility group 1 (HMGB1) and calreticulin (CRT)
were detected by immunofluorescence assay. As can be seen in Fig. 5i, strong red fluorescence overlapped with the cell nucleus in control and only laser groups, while that was diminished in
the Pd2Sn@GOx-SP and Pd2Sn@GOx-SP + laser groups, indicating that Pd2Sn@GOx-SP could trigger pyroptosis and release HMGB1 into the extracellular milieu. Meanwhile, more CRT expression was
detected on the cellular surface in the Pd2Sn@GOx-SP and Pd2Sn@GOx-SP + laser groups, whereas no significant signal in the control or only laser group (Fig. 5j). Moreover, the cells treated
with drugs showed significant adenosine triphosphate (ATP) release, wherein the leakage level of the Pd2Sn@GOx-SP + laser group was the highest compared with other groups (Supplementary Fig.
22e). All results clearly validated that Pd2Sn@GOx-SP could induce large-scale ICD effects mediated by pyroptosis. Furthermore, western blot analysis was also initiated to assess protein
expression during the pyroptosis process (Fig. 5k). In the pyroptotic pathway, NLRP3 inflammasome and Caspase-1 could be stimulated by abundant ROS and GOx, and subsequently achieve the
cleaving of GSDMD into N-GSDMD, which could lead to the perforation of the cellular membranes49. The high expression of NLRP3 could be observed in the group treated with Pd2Sn@GOx-SP due to
the two-pronged strategy. In addition, the Pd2Sn@GOx-SP-related group exhibited high expression of Cleaved Caspase-1 (C-Caspase-1) and N-GSDMD proteins, which could underpin the drilled
pores in cell membrane, leading to the secretion of damage-associated molecular patterns. To further confirm the GSDMD- or NLRP3-dependent pyroptosis induced by Pd2Sn@GOx-SP + laser, we have
knocked down the expression of GSDMD or NLRP3 in CT26 cells, then the cytotoxicity of Pd2Sn@GOx-SP + laser on the treated CT26 cells was measured. To begin with, transient transfection was
performed to establish GSDMD or NLRP3 knockdown CT26 cells, respectively. The transfection efficiency was verified by western blot analysis (Supplementary Fig. 22f). And the experimental
results verified that CT26 cells with low expression of GSDMD or NLRP3 had higher cell viability than the control group after being treated with Pd2Sn@GOx-SP + laser, respectively
(Supplementary Fig. 22g). Thus, the pyroptosis could be achieved by the treatment of Pd2Sn@GOx-SP + laser, which is dependent on the GSDMD and NLRP3. Considering the results in
immunofluorescence and western blot analysis, we attempted to offer in-depth evidence of pyroptosis induced by Pd2Sn@GOx-SP. In the most intuitive way, the cell morphology after various
treatments was recorded. As can be seen in Supplementary Fig. 22h, CT26 cells with the treatment of Pd2Sn@GOx-SP possessed more distinct swelling (bubbling) features compared with those in
the control group, indicating the occurrence of pyroptosis. Meanwhile, Bio-TEM was also performed to monitor the cells with the different treatments (Fig. 5l). It can be observed that the
cell membrane of the cells treated with Pd2Sn@GOx-SP exhibited extensive vacuolization and incomplete cell membranes, and mitochondria were deformed and swollen. These results provide
concrete evidence for Pd2Sn@GOx-SP-induced pyroptosis due to a large amount of ROS generation. Significantly, the tumor cells exhibited high levels of SLC7A11, which could import cystine to
promote GSH synthesis and resist pyroptosis50. Considering that we introduced GOx to constrict that process by consuming glucose, which could deplete the NADPH to prevent the reduction of
cystine, achieving the inhibition of GSH synthesis. The quantitative evaluation of intracellular glucose was conducted, the cells treated with GOx exhibited the obvious consumption of
glucose (Supplementary Fig. 22j). In addition, the aberrant buildup of intracellular cystine could lead to disulfide stress, which ultimately induces disulfidptosis (Fig. 5a). To begin with,
we performed non-reducing western blots to validate glucose starvation could induce disulfide-bond formation in the actin cytoskeleton proteins. As shown in Fig. 5m, the actin cytoskeleton
proteins (FLNA, TLN1, and MYH9) in CT26 cells exhibited slower migration on the smears following treatment with Pd2Sn@GOx-SP. Some of these proteins displayed exceptionally
high-molecular-weight bands near the stacking layer, whereas the control or laser-only groups did not show similar migration patterns, indicating that the actin cytoskeleton proteins formed
multiple intermolecular disulfide bonds under the treatment of Pd2Sn@GOx-SP. Subsequently, we evaluated the NADPH levels by performing various treatments on CT26 cells. As depicted in Fig.
5n, the cells handled with Pd2Sn@GOx-SP exhibited a significant increase in the NADP+/NADPH ratio (which indicates NADPH depletion) due to glucose starvation. In addition, the intracellular
GSH levels after various treatments were also assessed. After the treatment of Pd2Sn@GOx-SP, the GSH levels were obviously decreased (Supplementary Fig. 22k), which could enhance the
pyroptosis process. To further confirm the disulfidptosis induced by Pd2Sn@GOx-SP + laser, the dithiothreitol (DTT) as disulfidptosis inhibitor was added to the cells, and the cell viability
was analyzed by CCK-8 assay. The results showed that the cell viability of the cells added with DTT was obviously higher than that in the absence of DTT after being treated with
Pd2Sn@GOx-SP + laser (Supplementary Fig. 22l), demonstrating the disulfidptosis could be triggered by Pd2Sn@GOx-SP. Meanwhile, to confirm the critical role of SLC7A11-mediated cystine uptake
for disulfide stress, the CT26 cells were incubated with Pd2Sn@GOx-SP + laser after the treatment of SLC7A11 inhibitor (Erastin). Compared with that without the treatment of Erastin, the
cell death rate of CT26 cells exhibited an obvious decrease due to the inhibition of SLC7A11-mediated cystine uptake (Supplementary Fig. 22m), indicating that the disulfide stress originated
from the accumulation of cystine mediated by SLC7A11. In addition, the expression levels of SLC7A11 in multiple cancer cells (4T1, HeLa, SW1990, A549, and HepG2 cells) and normal cells
(L929 cells) were evaluated by western blot analysis (Supplementary Fig. 22i). The results showed the higher expression level of SLC7A11 in these cancer cells than that in normal cells,
leading to higher cell mortality in cancer cells compared to normal cells with low SLC7A11 expression. Besides that, the CT26 cells were treated with ferroptosis inhibitor (Ferrostatin-1) to
give insight into the role of ferroptosis in this work. The result showed that the CT26 cells treated with Pd2Sn@GOx-SP + laser also exhibited significant inhibitory effect after the
addition of Ferrostatin-1, which was similar to that without addition of Ferrostatin-1, demonstrating the effect of ferroptosis could be ignored (Supplementary Fig. 22n). Therefore,
GOx-induced disulfidptosis combined with Pd2Sn@GOx-SP-induced pyroptosis are the main manner for promoting tumor cell death. Furthermore, we also examined whether the Pd2Sn@GOx-SP + laser
treatment could alleviate cell migration by wound-healing assays (Supplementary Fig. 23a). Indeed, cell migration was largely inhibited after treatment with Pd2Sn@GOx-SP + laser for 12 and
24 h, while the control or only laser group had minimal effect on the cell migration (Supplementary Fig. 23b). Moreover, the transwell migration assay of CT26 cells after diverse treatments
was also performed to evaluate the migration ability. Similarly, the cells treated with Pd2Sn@GOx-SP + laser were greatly repressed (Supplementary Fig. 23c), illustrating that Pd2Sn@GOx-SP
had a significant inhibitory effect on cell migration. Therefore, GOx-induced disulfidptosis combined with Pd2Sn@GOx-SP-induced pyroptosis greatly promoted tumor cell death and inhibited its
migration. To confirm the role of pyroptosis on the augmenting of immunogenicity, the maturation degree of dendritic cells (DCs) was analyzed by flow cytometry analysis, which could improve
the antigen presentation ability and initiate the subsequent anti-tumor cascade immunity51,52. The results showed that the Pd2Sn@GOx-SP combined with laser irradiation can significantly
increase the proportion of CD80+CD86+ cells (Fig. 5o and Supplementary Fig. 23d), indicating that Pd2Sn@GOx-SP treated tumor cells could stimulate DCs activation and facilitate anti-tumor
immune responses. IN VIVO PA/CT IMAGING-GUIDED SYNERGISTIC ANTI-TUMOR THERAPY The results of in vitro cell experiments validated that the Pd2Sn@GOx-SP + laser irradiation treatment group
resulted in pyroptosis and disulfidptosis. Prior to evaluating the curative effect of the tumor in vivo, the PA/CT imaging capability was assessed based on the ideal light absorption and
high X-ray attenuation coefficient (Fig. 6a)53,54. To evaluate the potential of Pd2Sn@GOx-SP for PA imaging, Pd2Sn@GOx-SP was intravenously (i.v.) injected into CT26-bearing mice, and then
imaged at different time intervals by a Vevo LAZR-X system. As presented in Fig. 6b, the PA signals reached the strongest at 6 h post-injection, and subsequently weakened owing to the
elimination of Pd2Sn@GOx-SP from tumor tissues. Meanwhile, the biodistribution of Pd2Sn@GOx-SP in main organs was also detected by ex vivo PA imaging at 6 h post-injection (Supplementary
Fig. 24a), showing the highest accumulation of Pd2Sn@GOx-SP in the liver compared with other organs. In addition, the blood oxygen saturation (SaO2) in the tumor was monitored at various
times (Supplementary Fig. 24b). The results showed the efficient oxygen supply ability of Pd2Sn@GOx-SP for glucose consumption by GOx, which is beneficial to the in vivo cascade reaction
process. Similarly, the CT imaging ability of Pd2Sn@GOx-SP was also investigated both in vitro and in vivo. As presented in Supplementary Fig. 24c, the CT signal intensity gradually
strengthened as the concentration of Pd2Sn@GOx-SP increased. Moreover, the Hounsfield unit (HU) value was positively correlated to the concentration of Pd2Sn@GOx-SP with a slope of 21.75
(Supplementary Fig. 24d). Thereafter, the CT26-bearing mice were intratumorally (i.t.) and i.v. injected with Pd2Sn@GOx-SP to evaluate the CT imaging capability, respectively. Contrasted
with the control group, the tumor tissues of mice injected with Pd2Sn@GOx-SP presented a high CT density, suggesting an efficient CT imaging capacity (Fig. 6c, d). Furthermore, we performed
CT imaging on the same tumor-bearing mouse at different time points after being injected with Pd2Sn@GOx-SP. Within 6 h, the CT density in the tumor region of the mice i.v. injected with
Pd2Sn@GOx-SP were markedly enhanced with the injection time extension (Supplementary Fig. 24e), which showed the same trend as PA imaging, suggesting the efficient tumor accumulation of
Pd2Sn@GOx-SP. Besides, the real-time temperature change in the tumor site was also monitored by a thermal imaging camera after 6 h of Pd2Sn@GOx-SP injection. The temperature of the mice
tumor raised obviously under the irradiation of laser compared with that without injection of Pd2Sn@GOx-SP (Supplementary Fig. 24f), exhibiting the superior in vivo photothermal effect of
Pd2Sn@GOx-SP. To sum up, the Pd2Sn@GOx-SP possessed the potential for PA, CT, and infrared thermal imaging to guide tumor synergistic therapy. The biosafety and biocompatibility of
Pd2Sn@GOx-SP are prerequisites for in vivo treatment, thus the comprehensive evaluation was performed. To begin with, we examined the hemolysis induction of Pd2Sn@GOx-SP in red blood cells
(Supplementary Fig. 25). The hemolysis assay showed that no significant hemolysis appeared by Pd2Sn@GOx-SP, even if the concentration reached 500 μg/mL. Moreover, the biodistribution
assessment of Pd2Sn@GOx-SP i.v. injected into CT26-bearing mice was comprehensively analyzed by using ICP-OES. As displayed in Fig. 6e, the high contents of Pd were observed in the liver and
spleen, which were consistent with the results of ex vivo PA imaging. After 6 h of Pd2Sn@GOx-SP injection, the Pd levels reached a maximum concentration (9.07% ID g−1) in tumor regions, and
maintained a relatively high level (6.33% ID g−1) even at 24 h, demonstrating the superior tumor-homing efficiency of Pd2Sn@GOx-SP. The high biocompatibility of SP endowed Pd2Sn@GOx-SP with
stable blood circulation capacity. The blood circulation half-life time of Pd2Sn@GOx-SP was acquired, which was _t_1/2(α) = 0.15 h and _t_1/2(β) = 3.88 h (Fig. 6f). Simultaneously, as
presented in Fig. 6g, the elimination rate constant of Pd2Sn@GOx-SP in the first stage was obtained to be −0.5043 µg mL−1/h, which showed a decrease to −0.0279 µg mL−1/h after an interval of
1.86 h. Benefiting from the appropriate blood circulation, Pd2Sn@GOx-SP could achieve abundant accumulation in the tumor to exert an anti-tumor effect. Subsequently, the in vivo anti-cancer
efficacy of Pd2Sn@GOx-SP on CT26-bearing mice was further evaluated (Fig. 6h). Twenty-five female BALB/c mice established with the cancer model were randomly separated into five groups (_n_
= 5), containing control (G1), laser (G2), Pd2Sn-SP (G3), Pd2Sn@GOx-SP (G4), Pd2Sn@GOx-SP + laser (G5). All groups were i.v. administered with PBS or nano-drug at a dose of 10 mg kg−1, and
irradiated with laser after 6 h of injection. During the treatment process, no significant abnormal body weight fluctuation was displayed in the treated groups (Fig. 6i), indicating high
biosafety. As depicted in Fig. 6j, k, control or only laser irradiation resulted in rapid tumor growth, whereas Pd2Sn-SP alone moderately inhibited tumor growth. Comparatively, the
Pd2Sn@GOx-SP + laser group exerted a significant therapeutic effect with a suppression rate of 83.76% owing to the synergistic effect of pyroptosis with disulfidptosis. Moreover, the tumor
weight also exhibited distinct differences after various treatments. The mice in the group of control or only laser possessed heavier tumors while tumor weights of the mice treated with
nano-drug were reduced to varying degrees (Fig. 6l). Digital photos of the tumors dissociated from each mouse reflected the identical results (Fig. 6m). After that, the biocompatibility of
the Pd2Sn@GOx-SP was also evaluated by conducting the blood routine and blood biochemical analyses on the treated mice, which could provide the basis for potential practical applications of
malignancy treatments. No significant abnormalities were found in the liver and kidney function of the mice before and after i.v. administrated with Pd2Sn@GOx-SP (Fig. 6n). Moreover, there
were also no noticeable changes in hematological biomarkers contrasted with the control group, verifying the insignificant side effects of Pd2Sn@GOx-SP on hematological system. To further
validate the satisfactory anti-cancer effect of Pd2Sn@GOx-SP combined with laser, TdT-mediated dUTP nick-end labeling (TUNEL) and hematoxylin–eosin (H&E) staining were proceeded on the
slices of tumor tissue collected from the mice with various treatments (Fig. 6o). As expected, the highest degree of cell death was reflected in the group of Pd2Sn@GOx-SP + laser than the
other treatment groups, verifying a favorable tumor therapeutic effect of Pd2Sn@GOx-SP. Moreover, Ki67 staining was also performed and the group of Pd2Sn@GOx-SP + laser greatly decreased the
expression of Ki67 (Fig. 6o), indicating the prominent inhibition effect for tumor aggressiveness. Furthermore, the underlying therapeutic mechanism induced by Pd2Sn@GOx-SP via pyroptosis
was also confirmed through the immunohistochemical investigation of NLRP3, C-Caspase-1, and GSDMD. All of them were visibly elevated in the tumor tissues extracted from the mice treated with
Pd2Sn@GOx-SP + laser (Fig. 6o), evidencing the occurrence of pyroptosis in tumor cells. Significantly, the main organs of mice with diverse treatments showed no distinct inflammation or
pathological changes (Supplementary Fig. 26), which additionally confirmed the remarkable histocompatibility of Pd2Sn@GOx-SP. Furthermore, we constructed another tumor xenograft model using
4T1 mammary cancer cells to validate the excellent anti-tumor efficacy of Pd2Sn@GOx-SP (Supplementary Fig. 27a). The results demonstrated that the growth of 4T1 tumors was significantly
inhibited by the treatment of Pd2Sn@GOx-SP + laser (Supplementary Fig. 27b, c). Meanwhile, the body weight of mice in each treatment group exhibited no significant abnormality (Supplementary
Fig. 27d), confirming the biosafety of Pd2Sn@GOx-SP. The excellent tumor suppression effect could also be observed based on the H&E and TUNEL staining images of tumor tissue slices
(Supplementary Fig. 27e). Moreover, the H&E staining images of the major organs collected from the mice with diverse treatments showed no distinct inflammatory or pathological changes,
which further validated the excellent biosafety of Pd2Sn@GOx-SP (Supplementary Fig. 27f). These results highlighted the therapeutic potential of Pd2Sn@GOx-SP, which could induce both
pyroptosis and disulfidptosis, making it a promising candidate for treating various types of tumors. IN VIVO IMMUNE STIMULATION EFFECT The release of massive cytoplasmic contents induced by
tumor pyroptosis has been demonstrated to trigger immune responses for achieving anti-tumor immune activity and cancer immunotherapy (Fig. 7a)25. The reconstruction of the immunosuppressive
microenvironment elicited by Pd2Sn@GOx-SP was investigated by flow cytometry analysis. Similar to the in vitro experimental results, the Pd2Sn@GOx-SP + laser group induced a higher
proportion of DCs maturation in spleens than that of other groups (Fig. 7b, c). The mature DCs play an important role in antigen presentation, thereby activating the proliferation of naive T
cells and evoking an adaptive immune response44,55. To further confirm the immunotherapeutic effect of Pd2Sn@GOx-SP, the spleen infiltrating T cells were emphatically evaluated. As unfolded
in Fig. 7d, e, the proportions of CD4+ and CD8+ T cells in Pd2Sn@GOx-SP + laser group could reach 47.0% and 37.7%, which were over 1.9 and 4.4 folds more than those in the control group,
respectively. Importantly, the IFN-γ levels distinctly elevated in the tumors of the mice treated with Pd2Sn@GOx-SP + laser, which was 6.5 folds higher than the mice in control group (Fig.
7f, g), demonstrating the effective activation of CD8+ T cells56. Simultaneously, the activation of T cells in tumors was also analyzed (Supplementary Fig. 28a). The counts of CD8+ T cells
in the group of Pd2Sn@GOx-SP + laser increased by 4.3 folds than that in control group (Fig. 7h). Moreover, the expression level of granzyme B (GZMB) in CD8+ T cells is also a good marker of
anti-cancer immunity activation. The expression level of GZMB in tumors was evaluated by immunohistochemical analysis to confirm the activation of anti-cancer immunity. As can be seen in
Fig. 6o, the GZMB levels were significantly upregulated in the tumors of the mice treated with Pd2Sn@GOx-SP + laser compared with other groups, demonstrating the effective activation of CD8+
T cells to achieve anti-cancer immunity. Tumor-associated macrophages (TAMs) are the prominent immune cells present in the tumor stroma57,58. The activated macrophages mainly comprise
M1-type and M2-type, which show pro-inflammatory and anti-inflammatory properties, respectively. M2-type TAMs usually endow TME with characteristics of immunosuppression, promoting the
progression of tumor59,60. Consequently, the repolarization of M2-type TAMs to M1-type TAMs could reverse the tumor immunosuppression, which is beneficial for tumor therapeutic effect61,62.
Consequently, the capability of Pd2Sn@GOx-SP to alter the phenotype of TAMs was explored in mouse tumors (Supplementary Fig. 28b). M2-type TAMs obviously decreased in the Pd2Sn@GOx-SP +
laser group than that in control group, while the ratio of M1-type TAMs raised from 6.23% to 74.6% (Fig. 7i), indicating the laser-amplified and excellent tumor immunotherapy effect of
Pd2Sn@GOx-SP. All these data provide convincing evidence that the Pd2Sn@GOx-SP could recruit immune cells into tumors to induce specific anti-tumor immunity. Tumor immunotherapy could
stimulate the immune system to derive a long-term anti-tumor immunological response, which could prevent the recurrence and metastasis of tumors63. Encouraged by the excellent activation of
immunity and treatment effect on the primary tumor, we further investigated the efficacy of Pd2Sn@GOx-SP on mouse models with recurrence by a tumor-challenging assay (Fig. 7j). The body
weights and rechallenge tumor volumes of all the mice with various treatments were measured during the treatment process (Fig. 7k, l). The rechallenge tumors in the Pd2Sn@GOx-SP + laser
group were the smallest among all the groups (Fig. 7m), indicating that the immune memory induced by Pd2Sn@GOx-SP could effectively inhibit tumor recurrence. Subsequently, the immune
mechanism of the distal tumor tissues in various groups was further investigated. As depicted in Supplementary Fig. 29a, the ratio of CD8+ T cells in the distant tumors was increased to 17.3
after treatment with Pd2Sn@GOx-SP + laser, which was over 5.3 folds higher than that in control group. Simultaneously, the IFN-γ levels in distant tumors were also evaluated (Supplementary
Fig. 29b). Results indicated that the secretion of IFN-γ significantly increased in the Pd2Sn@GOx-SP treated group (Fig. 7n), which was beneficial to the subsequent activation of anti-tumor
immunity. Furthermore, depleting antibodies against CD8 (anti-CD8α) were injected into mice to deplete CD8+ T cells, and the tumor growth was monitored to confirm that the treatment involves
the activation of anti-cancer immunity (Supplementary Fig. 27a). After the depletion of CD8+ T cells by the administration of anti-CD8α, the therapeutic efficacy of Pd2Sn@GOx-SP + laser
decreased significantly (Supplementary Fig. 27g). All these data provide convincing evidence that Pd2Sn@GOx-SP could recruit immune cells to tumors, thereby inducing a specific anti-tumor
immune response. Motivated by satisfactory immune response of Pd2Sn@GOx-SP, the activation of immune memory was further evaluated by analyzing the proportion of effector memory T cells (TEM,
CD62L−CD44+) and central memory T cells (TCM, CD62L+CD44+) in spleens (Fig. 7o). The ratios of TEM in the group of Pd2Sn@GOx-SP + laser increased to 85.1%, which was higher than that in
control group, indicating a potent immune memory was formed. These results demonstrated that Pd2Sn@GOx-SP could effectively reshape the immunosuppressive TME and stimulate the intense immune
response, thereby effectively expunging dormant tumor cells and hindering their recurrence. After that, the inhibitory effects of Pd2Sn@GOx-SP on lung metastasis were further investigated
by establishing a pulmonary metastasis model to confirm the immune memory effect (Supplementary Fig. 30a). After all the treatments, the CT26-bearing mice were i.v. injected with CT26 cells
to imitate lung metastasis, and the lungs of the mice were harvested after 13 days. The photographs and H&E staining images of lung tissues showed that abundant nodules in the lung
collected from the control group, and a significant improvement was observed in the Pd2Sn@GOx-SP group, but there was no obvious metastasis in the Pd2Sn@GOx-SP + laser combined treatment
group (Supplementary Fig. 30b). In general, the Pd2Sn@GOx-SP with satisfactory biosecurity could promote the maturation and infiltration of cytotoxic T lymphocytes, which activated strong
systematic immune responses against tumor recurrence and metastasis. Therefore, the Pd2Sn@GOx-SP-mediated pyroptosis and disulfidptosis could effectively inhibit tumor growth, recurrence,
and spontaneous metastases. RNA-SEQUENCING ANALYSIS OF ANTI-CANCER MECHANISM Given the effective therapeutic effect and immune activation of Pd2Sn@GOx-SP on tumors, we further explored the
potential therapeutic mechanism by analyzing the mRNA profiles through high-throughput RNA sequencing. In total, 964 differentially expressed genes (DEGs) were screened out between the
Pd2Sn@GOx-SP + laser and control groups, of which 602 (62.45%) genes were upregulated and 362 (37.55%) genes were downregulated (Fig. 8a–c). Subsequently, the Kyoto Encyclopedia of Genes and
Genomes (KEGG) and Gene Ontology (GO) analysis of DEGs was conducted to examine the biological roles (Fig. 8d, e). The results suggested that the DEGs correlated with Pd2Sn@GOx-SP + laser
treatment were related to immunity pathways, demonstrating that Pd2Sn@GOx-SP + laser could activate the immune response by triggering programmed death of tumor cells. This discovery
highlights the importance of these genes in immune regulation, providing important clues for us to understand their potential roles in diseases or biological processes. In addition, the
correlation between the downregulated genes and cell adhesion was confirmed by the GO enrichment analysis, which is critical for cell migration, further confirming the inhibition of tumor
cell migration by the Pd2Sn@GOx-SP + laser treatment. Furthermore, gene set enrichment analysis (GSEA) confirmed that the DEGs were involved in pathways associated with pyroptosis (Fig. 8f).
Additionally, heat maps of DEGs associated with disulfidptosis were also generated (Fig. 8g). The expression of disulfidptosis-related genes (including NDUFA11, GYS1, OXSM, LRPPRC, NDUFS1,
NCKAP1, PRDX1, RPN1, and NUBPL) was notably upregulated in the Pd2Sn@GOx-SP + laser treatment group9. The evident genetic changes demonstrated that Pd2Sn@GOx-SP + laser could induce tumor
cell death via pyroptosis and disulfidptosis processes. DISCUSSION In summary, the precise pyroptosis and disulfidptosis dual-inducer of intermetallic Pd2Sn modified with GOx was developed
for exerting anti-tumor immune effects. Various morphologies (including NDs and NRs with different lengths) of Pd2Sn intermetallic compounds were constructed and the properties were further
contrasted. Both in vitro experiments and DFT calculations results confirmed that the NRs with the highest specific surface area exhibit the best catalytic performance. The intermetallic
Pd2Sn with ordered structure displayed enhanced NIR light absorption and multiple enzyme catalytic activities (CAT, POD, and OXD-mimic activity), which facilitate photothermal conversion and
ROS generation, thereby achieving GSDMD-dependent pyroptosis. Additionally, owing to the consumption of glucose by integrated GOx, Pd2Sn@GOx-SP could increase the NADP+/NADPH ratio, leading
to the abundant accumulation of cystine in CT26 cells with high SLC7A11 expression, achieving cystine-related disulfidptosis. Moreover, the enhanced cascade catalytic reaction could also be
realized by the generation of H2O2, photothermal effect, production of O2, and GSH depletion triggered by the complementary relationship of Pd2Sn and GOx. Notably, Pd2Sn@GOx-SP-induced
pyroptosis and disulfidptosis could effectively reprogram TME by alleviating immunosuppression and promoting T cell infiltration, thus promoting immune responses mediated by T cells, which
are conducive to the inhibition of tumor metastasis and recurrence. This work not only provided a strategy for the imaging-guided dual-inducer design of pyroptosis and disulfidptosis, but
also broadened the biomedical applications of intermetallic compounds, which offers good prospects for future advancement of cancer immunotherapy. METHODS ANIMAL CARE Female BALB/c mice
(4-week-old) were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China) (1100111084356). The animal experiments were conducted with the approval of ethics
by the Ethics Committee of Harbin Medical University Cancer Hospital (No. KY2024-33). Experimental group sizes were approved by the Regulatory Authorities for Animal Welfare after being
defined to balance statistical power, feasibility, and ethical aspects. CT26 cells or 4T1 cells (1 × 106, 100 μL in PBS) were subcutaneously injected into the right back of each BALB/c mouse
when they were 5 weeks old to establish CT26 or 4T1 tumor models, and the following experiments were conducted when the tumor volume approached 60 mm3. The maximal tumor size/burden
permitted by the ethics committee is 1500 mm3, and the maximal tumor size/burden was not exceeded in the experiments. The sex of the animal was not specifically considered in this study.
Mice of the appropriate sex were employed according to the requirements of the tumor model. A female mouse model was employed for CT26 colon cancer and 4T1 breast cancer. Mice were housed in
a specific-pathogen-free condition at 26 ± 1 °C and 50 ± 5% humidity with a 12-h light–dark cycle with unrestricted access to food and water. CHEMICALS AND MATERIALS Oleylamine, MAHC,
Pd(acac)2, Sn(OAc)2, TOP, GOx, SP, TMB, MB, DMPO, and OPD were purchased from Sigma-Aldrich (Shanghai, China). Ethanol and chloroform were of analytical grade and purchased from various
sources. MTT, DCFH-DA, 4′,6-diamidino-2-phenylindole (DAPI), Calcein-AM and propidium iodide (PI), JC-1 staining kit, and ActinGreen were purchased from Beyotime Inst. Biotech. (Haimen,
China). [Ru(dpp)3]Cl2 (cat: MX4826) was purchased from MKBIO (Shanghai, China). Anti-CRT (AF1666) and anti-HMGB1 (PH406) were purchased from Beyotime. ATP content assay kit was obtained from
Shanghai EnzymeLink Biotechnology Co., Ltd. (Shanghai China). The glutathione assay kit and glucose assay kit were purchased from Wanleibio Co., Ltd. Anti-Granzyme B (ab4059) was obtained
from Abcam. Filamin A Rabbit pAb (A0927) was purchased from Company ABclonal, Inc. MYH9 Polyclonal antibody (cat. 11128-1-AP) and Talin-1 Polyclonal antibody (cat. 14168-1-AP) were purchased
from Proteintech Group, Inc. Ferrostatin-1 and Erastin were obtained from MedChemExpress. DTT, RPMI 1640 medium, fetal bovine serum (FBS), and Dulbecco’s modified Eagle medium (DMEM) were
purchased from Thermo Fisher Scientific Inc. Anti-CD45-APC/Cyanine7 (cat.103116), anti-CD3-PerCP/Cyanine5.5 (cat.100218), anti-CD4-FITC (cat.100406), anti-CD8a-APC (cat.100712),
anti-CD11c-FITC (cat.117306), anti-CD86-APC (cat.105012), anti-CD80-PE (cat.104708), anti-CD11b-FITC (cat.101206), anti-F4/80-PerCP/Cyanine5.5 (cat.123126), anti-CD206-PE (cat.141706),
anti-CD44-FITC (cat.103022), anti-CD62L-PE (cat.161204), anti-IFN-γ-PE (cat.505808) were purchased from Biolegend (USA). InVivoMAb anti-mouse CD8α (cat. BE0061) was purchased from BioXCell.
H&E Stain Kit was obtained from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China). The TUNEL cell apoptosis detection kit was bought from Dalian Meilun Biotechnology
Co., Ltd. All chemicals were used as received without further treatment. CHARACTERIZATION The PXRD measurement was examined with a Rigaku D/max-TTR-III diffractometer using Cu-Ka radiation
(_λ_ = 0.15405 nm) at 40 kV and 40 mA. TEM and HRTEM were recorded on an FEI Tecnai G2 S-Twin transmission electron microscope equipped with a field emission gun operating at 200 kV. XPS
spectra were carried out using an ESCALAB 250 instrument. The zeta potential measurement for different samples was performed on a Malvern Zeta sizer Nan Nano ZS90. A UV-1601
spectrophotometer was used to obtain the UV–vis–NIR absorption spectrum. ESR spectra were acquired by a Bruker EMX1598 spectrometer. The element quantitative analysis of the sample was
conducted on ICP-OES (Agilent 725, Agilent Technologies, USA). The flow cytometry assays were performed on a BD Accuri C6 flow cytometer (USA). A CLSM (Leica TCS SP8) was adopted to obtain
the fluorescence image. SYNTHESIS OF PD2SN The Pd2Sn with different morphology were synthesized with the same procedure, except for the amount of MAHC added. Briefly, oleylamine (20 mL), a
certain quantity of MAHC, Pd(acac)2 (0.2 mmol), and Sn(OAc)2 (0.1 mmol) were added into the four-necked flask (100 mL). Under continuous stirring at 40 °C, the flask containing various
precursors was vacuumed for 20 min, then protected by injecting with nitrogen at 60 °C for 30 min to form a uniform solution. After that, the TOP (1 mL) was injected into a four-necked flask
and quickly heated up to 200 °C and maintained at this temperature for 30 min. Subsequently, the mixed solution in the flask was heated to 300 °C and preserved for another 30 min. Finally,
the flask was discontinued heating and cooled to room temperature (RT) naturally. Pd2Sn intermetallics were obtained by centrifugation and washed with ethanol and chloroform. In a
controllable way, the Pd2Sn NDs or NRs with different lengths were synthesized by adding MAHC with different amounts. SYNTHESIS OF PD2SN@GOX-SP The as-prepared Pd2Sn nanoparticles dissolved
in cyclohexane (1 mg mL−1, 10 mL) were added dropwise into chloroform solution of SP (2 mg mL−1, 30 mL). The solvent of the mixture was evaporated in a rotary evaporator under a vacuum at 60
°C to collect Pd2Sn-SP. Then, the obtained Pd2Sn-SP and GOx (5 mg) were dispersed in H2O and stirred vigorously overnight. The Pd2Sn@GOx-SP nanocomposites were performed by centrifugation
and washed with water and ethanol for further use. IN VITRO PHOTOTHERMAL PERFORMANCE OF PD2SN-SP The laser wavelength used in this work was 808 nm, which was generated by the 808 nm
multimode fiber coupled laser (Changchun Laser Optoelectronics Technology Co., Ltd). The power density of the 808 nm laser used in the in vitro or in vivo experiments was 0.8 W cm−2. The
rod-shaped Pd2Sn-SP solution with various concentrations was irradiated with laser at RT for 10 min and photographed at specified intervals by thermal imaging equipment (FLIR System E40). In
addition, pure water was performed as a comparison. Furthermore, the Pd2Sn-SP solution was irradiated with a laser for four cycles to explore the photothermal stability, and the change of
temperature was plotted. Selecting the cooling phase curve to calculate the photothermal conversion efficiency (_η_) of Pd2Sn-SP based on the following formula:
$$\eta=\frac{{hA}\left({T}_{max} -{T}_{surr}\right)-{Q}_{s}}{I(1-{10}^{-{A}_{\lambda }})}$$ (1) _h_, _A_, _T_max, _T_surr, _I_, and _A__λ_ refer to heat transfer coefficient, superficial
area, maximum equilibrium temperature, ambient temperature, laser fluence, and absorption value, respectively. _Q__s_ = (5.4 × 10−4) _I_. ESR MEASUREMENT The ·OH was measured by using DMPO
as the trapping agent through ESR analysis. The Pd2Sn-SP (100 μL, 1 mg mL−1), PBS (340 μL, pH = 5.5), and DMPO (10 μL) were mixed, and the solution was analyzed by an electron paramagnetic
resonance spectrometer after the rapid injection of H2O2 (100 mM, 50 μL). In addition, the stimulation of the laser was performed at the same reaction time, and the characteristic signal
peak with the intensity of 1:2:2:1 for ·OH was captured. CATALASE (CAT)-MIMIC ACTIVITY OF PD2SN-SP The detection of O2 generation induced by Pd2Sn-SP with different morphologies was
conducted to evaluate the CAT-mimic activity by using a dissolved oxygen meter. In a typical process, the Pd2Sn-SP (0, 50, 100, 200, and 400 μg mL−1) was mixed with H2O2, and the oxygen
concentration (mg L−1) was recorded last for 6 min, respectively. OXIDASE (OXD)-MIMIC ACTIVITY OF PD2SN-SP TMB as the substrate was used to measure the OXD-mimic activity of Pd2Sn-SP.
Typically, TMB (300 μg mL−1) and Pd2Sn-SP (200 μg mL−1) were dispersed in PBS (3 mL) at RT, and the absorbance was measured after a certain reaction time by using a UV–vis–NIR
spectrophotometer. GLUCOSE CONSUMPTION Pd2Sn@GOx-SP (0 and 200 μg mL−1) were mixed with glucose solution (1 mg mL−1). At various time intervals, the mixed solution (0.5 mL) was added with
dinitrosalicylic acid (DNS) reagent (1.5 mL). Then, the mixed solution was heated to 100 °C and maintained for 5 min, then cooled to RT. Subsequently, the absorbance of the mixed solution at
595 nm was measured by UV–vis–NIR spectrophotometer. PEROXIDASE (POD)-MIMIC ACTIVITY OF PD2SN-SP AND KINETIC ASSAY The POD-mimic activity for the ·OH generation of Pd2Sn-SP was measured
using TMB and OPD as the substrates. Briefly, 3 mL PBS including Pd2Sn-SP (100 μg mL−1) and TMB (300 μg mL−1) was mixed with H2O2, and the absorbance in various groups was recorded after a
certain reaction time. The groups exposed to a laser were performed at the same conditions except for laser action. The POD-mimicking kinetics of Pd2Sn-SP (NDs, short NRs, medium NRs, long
NRs) with different morphologies were also evaluated. TMB (300 μg mL−1), Pd2Sn-SP (100 μg mL−1), and H2O2 (final concentrations of 1, 2, 4, 8, and 16 mM) were mixed in PBS (3 mL) at RT, the
absorbance intensity at 652 nm were measured on a UV–vis–NIR spectrophotometer, and the Michaelis–Menten equation was adopted to analyze the Michaelis–Menten constant. The catalytic activity
of Pd2Sn-SP with different morphologies was further evaluated by altering the amounts of Pd2Sn-SP. Similarly, equivalent TMB, H2O2, and different amounts of Pd2Sn-SP were added into PBS (3
mL) at RT, and the absorbance of the mixed solution was recorded at various times. The catalytic activity (units) was calculated to make a comparison of the Pd2Sn-SP with different
morphologies. MB DEGRADATION ASSAY The prepared MB aqueous solution (10 μg mL−1) was mixed with Pd2Sn-SP to form a mixed solution uniformly, which was injected into the H2O2 solution (final
concentration of 50 μM) rapidly. The absorbance of the mixed solution was detected at various time intervals. DPA DEGRADATION ASSAY The generation of singlet oxygen (1O2) by Pd2Sn-SP was
measured using DPA as the substrate. First, DPA (1 mg mL−1) was dissolved in DMSO and mixed with Pd2Sn-SP. Then, an 808 nm laser was used to irradiate the mixture, the absorbance was
measured at various times. DENSITY FUNCTIONAL THEORY (DFT) CALCULATION The Vienna Ab initio Simulation Package was used for DFT calculation. The plane wave cutoff was set to 500 eV, and the
integration over the Brillouin zone was treated by the Monkhorst–Pack technique with a (2 × 2 × 1) grid for relevant surfaces and a (4 × 4 × 4) one for the bulk crystal. The energy and force
are converged when the values are <1 × 10−5 eV and −0.05 eV/Å, respectively. For bulk crystal, a (2 × 2 × 1) supercell containing 24 atoms was considered, while the (2 × 2 × 1) supercell
for the relevant Pd2Sn (001, 010, 011) surfaces was used. The (001, 010, 011) surfaces of different models were employed in the whole calculation process owing to the (001, 010, 011)
crystal planes are the main exposed crystal planes. CELL CULTURE The L929, 3T3, CT26, 4T1, HeLa, SW1990, A549, and HepG2 cells were obtained from the Heilongjiang Key Laboratory of Molecular
Oncology. The L929, HeLa, and HepG2 cells were seeded in minimum Eagle’s medium (Procell, Wuhan, China), 3T3 cells were seeded in DMEM (Procell, Wuhan, China), CT26 and 4T1 cells were
seeded in RPMI 1640 medium (Procell, Wuhan, China), SW1990 cells were seeded in Leibovitz’s L-15 medium (Procell, Wuhan, China), A549 cells were seeded in Ham’s F-12K medium (Procell, Wuhan,
China), and all medium were added with 10% FBS and 1% penicillin–streptomycin. The cells were cultured at 37 °C under a 5% CO2 atmosphere. CELLULAR UPTAKE AND SUBCELLULAR LOCALIZATION CT26
cells were seeded in 6-well plates and cultured for 24 h. Then, the cell culture medium was replaced with a fresh culture medium containing FITC-labeled Pd2Sn@GOx-SP for 0, 1, 2, 4, and 8 h,
respectively. For subcellular localization, the treated cells were further stained with commercial Lyso-Tracker Red DND-99 (100 nM) and DAPI according to the manufacturer’s guidelines. The
fluorescence images of cells were captured by CLSM after rinsing with PBS and fixing with glutaraldehyde (2.5%). The colocalization analysis was performed by Image J software. For the flow
cytometry assay, the treated cells were resuspended by trypsin and washed with PBS, and the fluorescence intensity of the cells was recorded by a flow cytometer. IN VITRO CYTOTOXICITY ASSAY
L929, 3T3, and CT26 cells were cultured in 96-well plates for 12 h. Then, the cell culture medium was refreshed with a fresh culture medium containing Pd2Sn-SP or Pd2Sn@GOx-SP with different
concentrations, and the laser-related groups were irradiated with an 808 nm laser. Finally, the standard MTT assay was conducted. Similarly, 4T1, HeLa, SW1990, A549, and HepG2 cells were
cultured in 96-well plates for 12 h. Then, these cells were treated with Pd2Sn@GOx-SP + laser, and the relative cell viability was obtained by MTT assay. IN VITRO CELL EXPERIMENTS EVALUATION
For most cell experiments, CT26 cells are subjected to the same culture and subsequent processing. Briefly, CT26 cells were seeded in a 6-well plate and cultured for 24 h. Then, the
cultivated cells were performed with different conditions, containing control (G1), laser (G2), Pd2Sn-SP (G3), Pd2Sn@GOx-SP (G4), Pd2Sn@GOx-SP + laser (G5). After co-incubation at 37 °C for
4 h, the laser-related groups were irradiated with 808 nm laser (0.8 W cm−2, 5 min), and then the cells were stained with the corresponding fluorescent dyes for analysis. For the detection
of intracellular ROS production, the treated cells were stained with DCFH-DA and DAPI for 20 and 15 min in the dark, respectively. Finally, the fluorescence of cells was captured by a CLSM.
For flow cytometry analysis, the treated cells were resuspended by trypsin and washed with PBS, and the fluorescence intensity of the cells was recorded by a flow cytometer. For the
detection of intracellular O2 generation, the cells were treated with [Ru(dpp)3]Cl2 (with the final concentration of 30 μM) for 6 h before various treatments. The treated cells were stained
with DAPI for 15 min. Finally, the fluorescence of cells was observed by a CLSM. For the detection of mitochondrial membrane potential, the treated cells were stained with JC-1 and DAPI for
20 and 15 min, respectively. Finally, the fluorescence of cells was detected by a CLSM. For the detection of living/dead cells, the treated cells were stained with Calcein-AM and PI at 37 °C
for 30 min. Finally, the fluorescence of cells was captured by a CLSM. For the cell death analysis, the treated cells were stained with Annexin V-FITC/PI apoptosis detection kit. Finally,
the cells were detected by a flow cytometer. For the observation of cell morphology, the morphology of the treated cells was directly observed by a fluorescence microscope. Furthermore, the
treated CT26 cells were treated with trypsin and then centrifuged to collect the precipitate, which was fixed, dehydrated, embedded, sliced, and measured by bio-TEM. For the CRT exposure and
HMGB1 migration analysis, the treated CT26 cells were washed with PBS and fixed with stationary liquid for 10 min. The anti-CRT and anti-HMGB1 were added to the cells and incubated at 4 °C
for overnight, respectively. Finally, the cells were further incubated by Alexa Fluor 488 or Alexa Fluor 555-conjugated secondary antibody and DAPI. Finally, the fluorescence of cells was
captured by a CLSM. IN VITRO TUMOR SPHEROID ASSAY CT26 cells were cultured in a 96-well plate coated with 1.5% agarose and monitored until the spheroid size reached about 600 μm. Then, fresh
culture medium containing FITC-labeled Pd2Sn@GOx-SP was adopted to replace the old culture medium and co-cultured for 0, 2, 4, and 8 h, respectively. Finally, the tumor spheroids were
cleaned with PBS and the fluorescence intensity was observed by a CLSM. For the cytotoxicity analysis of Pd2Sn@GOx-SP on tumor spheroids, the tumor spheroids were performed with different
conditions (G1, control; G2, laser; G3, Pd2Sn-SP; G4, Pd2Sn@GOx-SP; G5, Pd2Sn@GOx-SP + laser). After co-incubation at 37 °C for 8 h, the laser-related groups were exposed to laser, and then
the tumor spheroids were stained with Calcein-AM and PI for 30 min. Finally, the fluorescence of cells was captured by a CLSM. GSDMD AND NLRP3 KNOCKDOWN GSDMD and NLRP3 siRNAs obtained from
General Biology (Anhui, China) were transfected into CT26 cells. The siRNA sequences of GSDMD were (5′-CCGAGGUGCUGCAGACAAATT-3′) and (5′-UUUGUCUGCAGCACCUCGGTT-3′),
(5′-CCUCAGAACUGGAGAGCUUTT-3′) and (5′-AAGCUCUCCAGUUCUGAGGTT-3′), (5′-GCUAGAAGAAUGUGGCCUATT-3′) and (5′-UAGGCCACAUUCUUCUAGCTT-3′). The siRNA sequences for NLRP3 were
(5′-GAAAGAAACUGCUGCCCAATT-3′) and (5′-UUGGGCAGCAGUUUCUUUCTT-3′), (5′-GUACUUAAAUCGUGAAACATT-3′) and (5′-UGUUUCACGAUUUAAGUACTT-3′), (5′-GGAUGGGUUUGCUGGGAUATT-3′) and
(5′-UAUCCCAGCAAACCCAUCCTT-3′). Empty planter plasmid or siRNA control plasmid was used as a negative control. Briefly, CT26 cells were grown in 6-well plates and transformed with 2 μg of
plasmids using Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) according to the manufacturer’s protocol. After the transformation of 48 h, cells were harvested for further experiments. The
effectiveness of knockdown was confirmed by using western blot. WESTERN BLOTTING CT26 cells were seeded in a 6-well plate and cultured for 24 h. The cultivated cells were performed with
different conditions (G1–G5). Then, the cells were washed with cold PBS for twice and added to ice-cold lysis buffer for western blotting (Beyotime, China). Protein concentration was
determined using the BCA protein assay kit (Beyotime, China). The protein with the same amount was added to a 10% SDS polyacrylamide gel, electrophoresed, and transferred to polyvinylidene
fluoride (PVDF) membranes (Roche Diagnostics GmbH, Mannheim, Germany). Then, 5% skimmed milk was used to block the gels for 1 h, which were further incubated with primary antibodies and
secondary antibodies. Finally, the protein bands were detected with a fluorescent luminescence detector. The antibodies used in this study were as follows: NLRP3 (15101, CST), anti-GSDMD
antibody (69469, CST), anti-Caspase-1 antibody (3866, CST), anti-β-actin antibody (TA-09, ZSGB-BIO), anti-GSDMD-N (69469, CST), and SLC7A11 (PK30003, Proteintech). IN VITRO DENDRITIC CELLS
(DCS) STIMULATION The bone-marrow-derived DCs were obtained from the leg bones of male BALB/c mice. To begin with, the cultured CT26 cells were performed with different treatments (G1–G5).
Then, the treated CT26 cells were collected and co-cultured with immature DCs for 24 h. Finally, the non-adherent cells were collected and stained with anti-CD11c-FITC, anti-CD86-APC, and
anti-CD80-PE antibodies to analyze the maturation level of DCs by flow cytometry. IN VITRO AND IN VIVO PA AND CT IMAGING PERFORMANCE Pd2Sn-SP with various concentrations were prepared to
explore the in vitro imaging performance by the PA and CT equipment. As for the in vivo PA and CT imaging, the Pd2Sn-SP (100 μL) dissolved in saline was i.v. administrated into tumor-bearing
mice (at the dose of 10 mg kg−1). In addition, the Pd2Sn-SP (100 μL) dissolved in saline was also i.t. injected into tumor-bearing mice to further investigate the CT imaging effect. The PA
and CT imaging was obtained at various times by using a Vevo LAZR system (VisualSonics Inc. New York, NY) and a small animal X-ray CT imaging system (Quantum GX, PerkinElmer), respectively.
In addition, the blood oxygen saturation (SaO2) in the tumor was further monitored at various times. IN VIVO BIODISTRIBUTION AND PHARMACOKINETICS OF PD2SN@GOX-SP The tumor-bearing mice (_n_
= 3 mice for each group) were treated by i.v. injection with Pd2Sn@GOx-SP (at the dose of 10 mg kg−1). After 1, 3, 6, 12, and 24 h, the major organs and tumors were collected by sacrificing
these mice and analyzed by an ICP-OES. For pharmacokinetics analysis, the BALB/c mice (_n_ = 3 mice for each group) were treated by i.v. injection with Pd2Sn@GOx-SP (at the dose of 10 mg
kg−1). After 0, 2, 4, 8, 15, 30 min, 1, 2, 4, 8, 12, and 24 h, the blood was extracted and analyzed by an ICP-OES. IN VIVO THERAPEUTIC EVALUATION OF PD2SN@GOX-SP The tumor-bearing mice were
randomly allocated into five groups (_n_ = 5 mice for each group), including control (G1), laser (G2), Pd2Sn-SP (G3), Pd2Sn@GOx-SP (G4), Pd2Sn@GOx-SP + laser (G5). The mice were i.v.
injected with saline or nano-drug on days 1, 5, 9, and 13. During the treatment process, the body weight and tumor size of the mice were measured every 2 days. The tumor volume (_V_) was
calculated according to _V_ = _lw_2/2. The mice were sacrificed on day 15, and the tumors and main organs were collected for further analysis. The tumor sections of different groups were
stained with H&E, TUNEL, Ki67, C-Caspase-1, and NLRP3. The main organ sections were stained with H&E. To conduct the tumor rechallenge assay, the tumor-bearing mice were randomly
allocated into five groups (G1–G5, _n_ = 5 mice for each group). On day 12, the mice were rechallenged with CT26 cells (1 × 106) on the left back. The body weight and tumor size were
measured every 2 days. On day 26, the mice were sacrificed and the spleens were collected to detect immune cells. To explore the in vivo anti-metastasis efficacy, the tumor-bearing mice were
randomly allocated into five groups (G1–G5, _n_ = 3 mice for each group). After being treated for 12 d, the CT26 cells were i.v. injected into these mice. On day 25, the mice were all
sacrificed to collect the lungs for H&E staining assays. IN VIVO ANTI-TUMOR IMMUNITY For the macrophage polarization analysis, the tumors of mice with different treatments were
extracted, homogenized in PBS, and filtered to obtain single-cell suspension. Subsequently, the cells were stained with anti-CD45-APC/Cyanine7, anti-CD11b-FITC, anti-F4/80-PerCP/Cyanine5.5,
anti-CD86-APC, and anti-CD206-PE antibodies, and the macrophage phenotype was analyzed by flow cytometry. For T-cell activation analysis, the collected spleens and tumors were homogenized in
PBS, and filtered to obtain single-cell suspension. Afterwards, the cells were stained with anti-CD45-APC/Cyanine7, anti-CD3-PerCP/Cyanine5.5, anti-CD4-FITC, and anti-CD8a-APC, and the
activation of T cells was analyzed by flow cytometry. The activated CD8 T cells were further analyzed by staining primary and distant tumor cells with anti-IFN-γ-PE antibodies. For DCs
maturation analysis, the collected spleens were homogenized in PBS, and filtered to obtain single-cell suspension. Afterwards, the cells were stained with anti-CD11c-FITC, anti-CD86-APC, and
anti-CD80-PE antibodies to analyze the maturation of DCs using flow cytometry. To analyze memory T cells, the collected spleens were homogenized in PBS, and filtered to obtain single-cell
suspension. Afterwards, the cells were stained with anti-CD45-APC/Cyanine7, anti-CD3-PerCP/Cyanine5.5, anti-CD8a-APC, anti-CD44-FITC, anti-CD62L-PE antibodies to evaluate the memory T cell
using flow cytometry. RNA SEQUENCING The tumor-bearing mice were randomly allocated into two groups, including control and Pd2Sn@GOx-SP + laser. After 1 week of treatment, the mice were
sacrificed and the tumors were extracted for RNA sequencing according to the requirement of GENEWIZ, Inc. (Suzhou, China). STATISTICAL ANALYSIS Quantitative data were indicated as mean ±
S.D. The software of GraphPad Prism 9.0 was adopted to assess the statistical analysis, which was performed using the Student’s _t_-test and one/two-way ANOVA. The statistical significance
was attained at _*p_ < 0.05, _**p_ < 0.01, _***p_ < 0.001, and _****p_ < 0.0001. REPORTING SUMMARY Further information on research design is available in the Nature Portfolio
Reporting Summary linked to this article. DATA AVAILABILITY The RNA-seq data generated in this study are available in the GSA database under accession code CRA016541 and could be achieved by
the following hyperlink [https://ngdc.cncb.ac.cn/gsa/browse/CRA016541]. The remaining data generated in this study are available within the Article, Supplementary Information, or Source
Data file. Source data are provided with this paper. REFERENCES * Siegel, R. L., Giaquinto, A. N. & Jemal, A. Cancer statistics, 2024. _CA Cancer J. Clin._ 74, 12–49 (2024). Article
PubMed Google Scholar * Dizon, D. S. & Kamal, A. H. Cancer statistics 2024: all hands on deck. _CA Cancer J. Clin._ 74, 8–9 (2024). Article PubMed Google Scholar * Santucci, C. et
al. European cancer mortality predictions for the year 2024 with focus on colorectal cancer. _Ann. Oncol._ 35, 308–316 (2024). Article PubMed CAS Google Scholar * Zhang, W., Hu, X.,
Shen, Q. & Xing, D. Mitochondria-specific drug release and reactive oxygen species burst induced by polyprodrug nanoreactors can enhance chemotherapy. _Nat. Commun._ 10, 1704 (2019).
Article ADS PubMed PubMed Central Google Scholar * Malla, J. A. et al. A glutathione activatable ion channel induces apoptosis in cancer cells by depleting intracellular glutathione
levels. _Angew. Chem. Int. Ed._ 59, 7944–7952 (2020). Article ADS CAS Google Scholar * Machesky, L. M. Deadly actin collapse by disulfidptosis. _Nat. Cell Biol._ 25, 375–376 (2023).
Article PubMed CAS Google Scholar * Liu, X., Zhuang, L. & Gan, B. Disulfidptosis: disulfide stress-induced cell death. _Trends Cell Biol._ 34, 327–337 (2023). Article PubMed Google
Scholar * Liu, X. et al. Cystine transporter regulation of pentose phosphate pathway dependency and disulfide stress exposes a targetable metabolic vulnerability in cancer. _Nat. Cell
Biol._ 22, 476–486 (2020). Article PubMed PubMed Central CAS Google Scholar * Liu, X. et al. Actin cytoskeleton vulnerability to disulfide stress mediates disulfidptosis. _Nat. Cell
Biol._ 25, 404–414 (2023). Article PubMed PubMed Central CAS Google Scholar * Koppula, P., Zhuang, L. & Gan, B. Cystine transporter SLC7A11/xCT in cancer: ferroptosis, nutrient
dependency, and cancer therapy. _Protein Cell_ 12, 599–620 (2020). Article PubMed PubMed Central Google Scholar * Yan, Y. et al. SLC7A11 expression level dictates differential responses
to oxidative stress in cancer cells. _Nat. Commun._ 14, 3673 (2023). Article ADS PubMed PubMed Central CAS Google Scholar * Feng, Q. et al. Severely polarized extracellular acidity
around tumour cells. _Nat. Biomed. Eng_. 2157, 1–13 (2024). * Lin, F., Li, M., Zeng, L., Luo, M. & Guo, S. Intermetallic nanocrystals for fuel-cells-based electrocatalysis. _Chem. Rev._
123, 12507–12593 (2023). Article PubMed CAS Google Scholar * Zhou, M., Li, C. & Fang, J. Noble-metal based random alloy and intermetallic nanocrystals: syntheses and applications.
_Chem. Rev._ 121, 736–795 (2021). Article PubMed CAS Google Scholar * Yan, Y. et al. Intermetallic nanocrystals: syntheses and catalytic applications. _Adv. Mater._ 29, 1605997 (2017).
Article Google Scholar * Cui, Z. et al. Synthesis of structurally ordered Pt3Ti and Pt3V nanoparticles as methanol oxidation catalysts. _J. Am. Chem. Soc._ 136, 10206–10209 (2014). Article
PubMed CAS Google Scholar * Gamler, J. T. L., Ashberry, H. M., Skrabalak, S. E. & Koczkur, K. M. Random alloyed versus intermetallic nanoparticles: a comparison of electrocatalytic
performance. _Adv. Mater._ 30, 1801563 (2018). Article Google Scholar * Chou, N. H. & Schaak, R. E. Shape-controlled conversion of β-Sn nanocrystals into intermetallic M-Sn (M = Fe,
Co, Ni, Pd) nanocrystals. _J. Am. Chem. Soc._ 129, 7339–7345 (2007). Article PubMed CAS Google Scholar * Zhu, P. et al. MnOOH-catalyzed autoxidation of glutathione for reactive oxygen
species production and nanocatalytic tumor innate immunotherapy. _J. Am. Chem. Soc._ 145, 5803–5815 (2023). Article PubMed CAS Google Scholar * He, S. et al. A semiconducting
iron-chelating nano-immunomodulator for specific and sensitized sono-metallo-immunotherapy of cancer. _Angew. Chem. Int. Ed._ 62, e202310178 (2023). Article CAS Google Scholar * Wang, W.
et al. A self-adaptive pyroptosis inducer: optimizing the catalytic microenvironment of nanozymes by membrane-adhered microbe enables potent cancer immunotherapy. _Adv. Mater._ 36, 2310063
(2024). Article CAS Google Scholar * Xu, X. et al. Virus-like particle-induced cGAS-STING activation and AIM2 inflammasome-mediated pyroptosis for robust cancer immunotherapy. _Angew.
Chem. Int. Ed._ 62, e202303010 (2023). Article CAS Google Scholar * Li, F. et al. mRNA lipid nanoparticle-mediated pyroptosis sensitizes immunologically cold tumors to checkpoint
immunotherapy. _Nat. Commun._ 14, 4223 (2023). Article ADS PubMed PubMed Central CAS Google Scholar * Niu, R. et al. Programmed targeting pyruvate metabolism therapy amplified
single-atom nanozyme-activated pyroptosis for immunotherapy. _Adv. Mater._ 36, 2312124 (2024). Article CAS Google Scholar * Wang, Q. et al. A bioorthogonal system reveals antitumour
immune function of pyroptosis. _Nature_ 579, 421–426 (2020). Article ADS PubMed CAS Google Scholar * Yu, B. et al. Platinum prodrug nanoparticles with COX-2 inhibition amplify
pyroptosis for enhanced chemotherapy and immune activation of pancreatic cancer. _Adv. Mater._ 36, 2310456 (2024). Article CAS Google Scholar * Zhao, Q. et al. Dual active centers linked
by a reversible electron station as a multifunctional nanozyme to induce synergetically enhanced cascade catalysis for tumor-specific therapy. _J. Am. Chem. Soc._ 145, 12586 (2023). Article
PubMed CAS Google Scholar * Lin, H., Wang, Y., Gao, S., Chen, Y. & Shi, J. Theranostic 2D tantalum carbide (MXene). _Adv. Mater._ 30, 1703284 (2018). Article Google Scholar * Liu,
P. et al. Photochemical route for synthesizing atomically dispersed palladium catalysts. _Science_ 352, 797 (2016). Article ADS PubMed CAS Google Scholar * Huang, L. et al. Synergy
between plasmonic and electrocatalytic activation of methanol oxidation on palladium-silver alloy nanotubes. _Angew. Chem. Int. Ed._ 58, 8794–8798 (2019). Article CAS Google Scholar * Li,
S. et al. Degradable holey palladium nanosheets with highly active 1D nanoholes for synergetic phototherapy of hypoxic tumors. _J. Am. Chem. Soc._ 142, 5649–5656 (2020). Article PubMed
CAS Google Scholar * Li, S. et al. Tensile-strained palladium nanosheets for synthetic catalytic therapy and phototherapy. _Adv. Mater._ 34, 2202609 (2022). Article CAS Google Scholar *
Fu, Q., Zhu, R., Song, J., Yang, H. & Chen, X. Photoacoustic imaging: contrast agents and their biomedical applications. _Adv. Mater._ 31, 1805875 (2019). Article Google Scholar *
Lin, L. & Wang, L. V. The emerging role of photoacoustic imaging in clinical oncology. _Nat. Rev. Clin. Oncol._ 19, 365–384 (2022). Article PubMed Google Scholar * Jia, X., Wang, J.
& Wang, E. Rapid synthesis of trimetallic nanozyme for sustainable cascaded catalytic therapy via tumor microenvironment remodulation. _Adv. Mater._ 36, 2309261 (2024). Article CAS
Google Scholar * Wang, Z. et al. 2D catalytic nanozyme enables cascade enzyodynamic effect-boosted and Ca2+ overload-induced synergistic ferroptosis/apoptosis in tumor. _Adv. Mater_. 36,
2312316 (2024). * Xu, J. et al. Carbon dioxide electroreduction into syngas boosted by a partially delocalized charge in molybdenum sulfide selenide alloy monolayers. _Angew. Chem. Int. Ed._
56, 9121–9125 (2017). Article ADS CAS Google Scholar * Sousa de Almeida, M. et al. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. _Chem. Soc.
Rev._ 50, 5397–5434 (2021). Article PubMed PubMed Central CAS Google Scholar * Donahue, N. D., Acar, H. & Wilhelm, S. Concepts of nanoparticle cellular uptake, intracellular
trafficking, and kinetics in nanomedicine. _Adv. Drug Deliv. Rev._ 143, 68–96 (2019). Article PubMed CAS Google Scholar * Wang, Y.-F. et al. Spatiotemporal tracing of the cellular
internalization process of rod-shaped nanostructures. _ACS Nano_ 16, 4059–4071 (2022). Article PubMed CAS Google Scholar * Rennick, J. J., Johnston, A. P. R. & Parton, R. G. Key
principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. _Nat. Nanotechnol._ 16, 266–276 (2021). Article ADS PubMed CAS Google Scholar * Yao, C.
et al. Near-infrared upconversion mesoporous cerium oxide hollow biophotocatalyst for concurrent pH-/H2O2-responsive O2-evolving synergetic cancer therapy. _Adv. Mater._ 30, 1704833 (2018).
Article Google Scholar * Yu, P. et al. Pyroptosis: mechanisms and diseases. _Signal Transduct. Target. Ther._ 6, 128 (2021). Article PubMed PubMed Central Google Scholar * Zhang, L. et
al. Integration of AIEgens into covalent organic frameworks for pyroptosis and ferroptosis primed cancer immunotherapy. _Nat. Commun._ 14, 5355 (2023). Article ADS PubMed PubMed Central
CAS Google Scholar * Kalyane, D. et al. Reactive oxygen nano-generators for cancer therapy. _Prog. Mater. Sci._ 130, 100974 (2022). Article CAS Google Scholar * Chang, M. et al.
Ultrasound-amplified enzyodynamic tumor therapy by perovskite nanoenzyme-enabled cell pyroptosis and cascade catalysis. _Adv. Mater._ 35, 2208817 (2023). Article CAS Google Scholar *
Yang, Y. et al. Electrostatic attractive self-delivery of siRNA and light-induced self-escape for synergistic gene therapy. _Adv. Mater._ 35, 2301409 (2023). Article CAS Google Scholar *
Xu, K. et al. Multienzyme-mimicking LaCoO3 nanotrigger for programming cancer-cell pyroptosis. _Adv. Mater._ 35, 2302961 (2023). Article CAS Google Scholar * Zhang, S. et al.
Biomineralized two-enzyme nanoparticles regulate tumor glycometabolism inducing tumor cell pyroptosis and robust antitumor immunotherapy. _Adv. Mater._ 34, 2206851 (2022). Article CAS
Google Scholar * Aboushousha, R. et al. Glutaredoxin attenuates glutathione levels _via_ deglutathionylation of Otub1 and subsequent destabilization of system xC−. _Sci. Adv._ 9, eadi5192
(2023). Article PubMed PubMed Central CAS Google Scholar * Zhao, X. et al. Orchestrated yolk–shell nanohybrids regulate macrophage polarization and dendritic cell maturation for
oncotherapy with augmented antitumor immunity. _Adv. Mater._ 34, 2108263 (2022). Article CAS Google Scholar * Palucka, K. & Banchereau, J. Cancer immunotherapy _via_ dendritic cells.
_Nat. Rev. Cancer_ 12, 265–277 (2012). Article PubMed PubMed Central CAS Google Scholar * Liu, Y. et al. Chemical design of activatable photoacoustic probes for precise biomedical
applications. _Chem. Rev._ 122, 6850–6918 (2022). Article PubMed CAS Google Scholar * Lee, N., Choi, S. H. & Hyeon, T. Nano-sized CT contrast agents. _Adv. Mater._ 25, 2641–2660
(2013). Article PubMed CAS Google Scholar * Zhang, L. et al. Engineering multienzyme-mimicking covalent organic frameworks as pyroptosis inducers for boosting antitumor immunity. _Adv.
Mater._ 34, 2108174 (2022). Article CAS Google Scholar * Hoekstra, M. E. et al. Long-distance modulation of bystander tumor cells by CD8+ T-cell-secreted IFN-γ. _Nat. Cancer_ 1, 291–301
(2020). Article PubMed PubMed Central CAS Google Scholar * Sylvestre, M., Crane, C. A. & Pun, S. H. Progress on modulating tumor-associated macrophages with biomaterials. _Adv.
Mater._ 32, 1902007 (2020). Article CAS Google Scholar * DeNardo, D. G. & Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. _Nat. Rev. Immunol._ 19, 369–382
(2019). Article PubMed PubMed Central CAS Google Scholar * Chen, S. et al. Macrophages in immunoregulation and therapeutics. _Signal Transduct. Target. Ther._ 8, 207 (2023). Article
ADS PubMed PubMed Central Google Scholar * Xia, Y. et al. Engineering macrophages for cancer immunotherapy and drug delivery. _Adv. Mater._ 32, 2002054 (2020). Article CAS Google
Scholar * Ovais, M., Guo, M. & Chen, C. Tailoring nanomaterials for targeting tumor-associated macrophages. _Adv. Mater._ 31, 1808303 (2019). Article Google Scholar * Zhang, X. et al.
Reprogramming tumour-associated macrophages to outcompete cancer cells. _Nature_ 619, 616–623 (2023). Article ADS PubMed PubMed Central CAS Google Scholar * Guo, Y., Hu, P. & Shi,
J. Nanomedicine remodels tumor microenvironment for solid tumor immunotherapy. _J. Am. Chem. Soc._ 146, 10217–10233 (2024). Article PubMed CAS Google Scholar Download references
ACKNOWLEDGEMENTS Financial support from the National Natural Science Foundation of China (NSFC 52002091 to L.F., U22A20347 to P.Y., and U22A20316 to Y. Zhou), Heilongjiang Natural Science
Foundation Project of Outstanding Youth Project (YQ2023B005 to L.F.), Natural Science Foundation of Heilongjiang Provincial Government (LH2022H067 to B.L.), China Postdoctoral Science
Foundation (2023TQ0091 to L.F.), Heilongjiang Postdoctoral Fund (LBH-Z23014 to B.L.), Ph.D. Student Research and Innovation Fund of the Fundamental Research Funds for the Central
Universities (3072023GIP1002 to Y. Zhu), and the Fundamental Research Funds for the Central Universities. The authors would like to thank Ruifang Shen from Harbin Institute of Technology,
Institute of Space Environment and Materials Science (National Major Science Engineering) for CT and PA imaging tests. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Key Laboratory of
Superlight Materials and Surface Technology, Ministry of Education, College of Materials Science and Chemical Engineering, Harbin Engineering University, Harbin, PR China Yanlin Zhu, Lili
Feng, Ruoxi Zhao, Bin Liu & Piaoping Yang * Department of Radiology, Harbin Medical University Cancer Hospital, Harbin, PR China Xinxin Wang, Can Yu & Yang Zhou * Department of
Biochemistry and Molecular Biology, Harbin Medical University, Harbin, PR China Yuanli Liu * Key Laboratory of Functional Inorganic Material Chemistry, Ministry of Education, School of
Chemistry and Materials Science, Heilongjiang University, Harbin, PR China Ying Xie Authors * Yanlin Zhu View author publications You can also search for this author inPubMed Google Scholar
* Xinxin Wang View author publications You can also search for this author inPubMed Google Scholar * Lili Feng View author publications You can also search for this author inPubMed Google
Scholar * Ruoxi Zhao View author publications You can also search for this author inPubMed Google Scholar * Can Yu View author publications You can also search for this author inPubMed
Google Scholar * Yuanli Liu View author publications You can also search for this author inPubMed Google Scholar * Ying Xie View author publications You can also search for this author
inPubMed Google Scholar * Bin Liu View author publications You can also search for this author inPubMed Google Scholar * Yang Zhou View author publications You can also search for this
author inPubMed Google Scholar * Piaoping Yang View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS These authors contributed equally: Y. Zhu
and X. Wang. Y. Zhu and X.W. conceived the idea and designed the project. Y. Zhu, X.W., R.Z., and C.Y. performed the experiments and analyzed the results. L.F., Y.L., B.L., and Y. Zhou
assisted with the experiment design and data analysis. Y. Zhu wrote and revised the original draft of the manuscript. Y.X. performed the DFT calculations. L.F., Y.X., and Y. Zhou reviewed
and edited the manuscript. P.Y. supervised the whole project. All authors discussed the results and commented on the manuscript. CORRESPONDING AUTHORS Correspondence to Lili Feng, Yang Zhou
or Piaoping Yang. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Communications_ thanks Jie Han, Kohsuke
Tsuchiya, XueBo Yin, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available. ADDITIONAL INFORMATION PUBLISHER’S NOTE
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION PEER REVIEW FILE
REPORTING SUMMARY SOURCE DATA SOURCE DATA RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International
License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the
source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived
from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line
to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will
need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/. Reprints and permissions ABOUT THIS
ARTICLE CITE THIS ARTICLE Zhu, Y., Wang, X., Feng, L. _et al._ Intermetallics triggering pyroptosis and disulfidptosis in cancer cells promote anti-tumor immunity. _Nat Commun_ 15, 8696
(2024). https://doi.org/10.1038/s41467-024-53135-2 Download citation * Received: 12 May 2024 * Accepted: 03 October 2024 * Published: 08 October 2024 * DOI:
https://doi.org/10.1038/s41467-024-53135-2 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative