Play all audios:
ABSTRACT Haematopoietic ageing is marked by a loss of regenerative capacity and skewed differentiation from haematopoietic stem cells (HSCs), leading to impaired blood production. Signals
from the bone marrow niche tailor blood production, but the contribution of the old niche to haematopoietic ageing remains unclear. Here we characterize the inflammatory milieu that drives
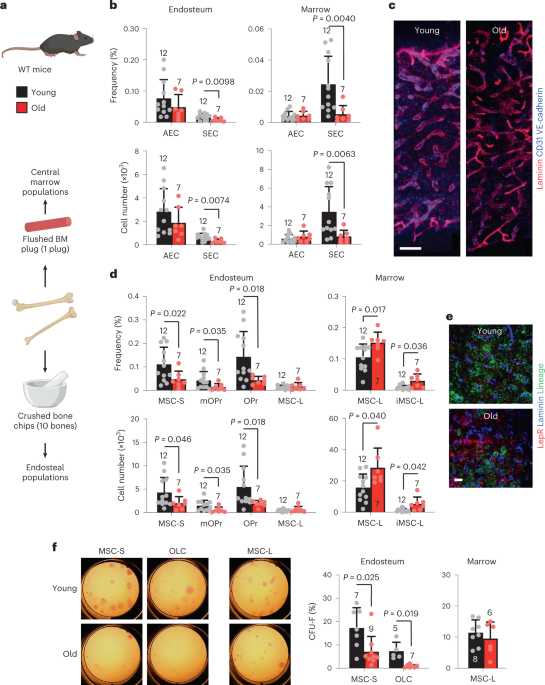
both niche and haematopoietic remodelling. We find decreased numbers and functionality of osteoprogenitors at the endosteum and expansion of central marrow LepR+ mesenchymal stromal cells
associated with deterioration of the sinusoidal vasculature. Together, they create a degraded and inflamed old bone marrow niche. Niche inflammation in turn drives the chronic activation of
emergency myelopoiesis pathways in old HSCs and multipotent progenitors, which promotes myeloid differentiation and hinders haematopoietic regeneration. Moreover, we show how production of
interleukin-1β (IL-1β) by the damaged endosteum acts in _trans_ to drive the proinflammatory nature of the central marrow, with damaging consequences for the old blood system. Notably, niche
deterioration, HSC dysfunction and defective regeneration can all be ameliorated by blocking IL-1 signalling. Our results demonstrate that targeting IL-1 as a key mediator of niche
inflammation is a tractable strategy to improve blood production during ageing. Access through your institution Buy or subscribe This is a preview of subscription content, access via your
institution ACCESS OPTIONS Access through your institution Access Nature and 54 other Nature Portfolio journals Get Nature+, our best-value online-access subscription $29.99 / 30 days cancel
any time Learn more Subscribe to this journal Receive 12 print issues and online access $209.00 per year only $17.42 per issue Learn more Buy this article * Purchase on SpringerLink *
Instant access to full article PDF Buy now Prices may be subject to local taxes which are calculated during checkout ADDITIONAL ACCESS OPTIONS: * Log in * Learn about institutional
subscriptions * Read our FAQs * Contact customer support SIMILAR CONTENT BEING VIEWED BY OTHERS MICRO-ENVIRONMENTAL SENSING BY BONE MARROW STROMA IDENTIFIES IL-6 AND TGFΒ1 AS REGULATORS OF
HEMATOPOIETIC AGEING Article Open access 14 August 2020 THE ROLE OF THE HAEMATOPOIETIC STEM CELL NICHE IN DEVELOPMENT AND AGEING Article 10 September 2024 NICHE DERIVED NETRIN-1 REGULATES
HEMATOPOIETIC STEM CELL DORMANCY VIA ITS RECEPTOR NEOGENIN-1 Article Open access 27 January 2021 DATA AVAILABILITY The RNA-seq and microarray data that support the findings of this study
have been deposited in the Gene Expression Omnibus under accession code GSE169162. The _M._ _musculus_ genome GRCm38.p4 is available from the National Center for Biotechnology Information
(https://www.ncbi.nlm.nih.gov/assembly/GCF_000001635.24/). The chip annotation MSigDB.v7.2.chip is available from the Broad Institute
(https://software.broadinstitute.org/cancer/software/gsea/wiki/index.php/MSigDB_v7.2_Release_Notes). Gene sets used for GSEA h.all.v7.2.symbols.gmt and c2.cp.reactome.v7.2.symbols.gmt are
available from the Broad Institute (https://data.broadinstitute.org/gsea-msigdb/msigdb/release/7.2/). All other data are available from the corresponding author upon reasonable request.
Source data are provided with this paper. CODE AVAILABILITY All code and packages used to support the findings of this study are either publicly available or available from the corresponding
author upon reasonable request. REFERENCES * Rossi, D. J., Jamieson, C. H. M. & Weissman, I. L. Stems cells and the pathways to aging and cancer. _Cell_ 132, 681–696 (2008). Article
CAS Google Scholar * Morrison, S. J. & Scadden, D. T. The bone marrow niche for haematopoietic stem cells. _Nature_ 505, 327–334 (2014). Article CAS Google Scholar * Pinho, S. &
Frenette, P. S. Haematopoietic stem cell activity and interactions with the niche. _Nat. Rev. Mol. Cell Biol._ 20, 303–320 (2019). Article CAS Google Scholar * Verovskaya, E. V.,
Dellorusso, P. V. & Passegué, E. Losing sense of self and surroundings: hematopoietic stem cell aging and leukemic transformation. _Trends Mol. Med._ 25, 494–515 (2019). Article Google
Scholar * Ho, Y.-H. & Méndez-Ferrer, S. Microenvironmental contributions to hematopoietic stem cell aging. _Haematologica_ 105, 38–46 (2020). Article CAS Google Scholar * Pietras, E.
M. et al. Functionally distinct subsets of lineage-biased multipotent progenitors control blood production in normal and regenerative conditions. _Cell Stem Cell_ 17, 35–46 (2015). Article
CAS Google Scholar * Olson, O. C., Kang, Y.-A. & Passegué, E. Normal hematopoiesis is a balancing act of self-renewal and regeneration. _Cold Spring Harb. Perspect. Med._ 10, a035519
(2020). Article CAS Google Scholar * Schepers, K., Campbell, T. B. & Passegué, E. Normal and leukemic stem cell niches: insights and therapeutic opportunities. _Cell Stem Cell_ 16,
254–267 (2015). Article CAS Google Scholar * Kovtonyuk, L. V., Fritsch, K., Feng, X., Manz, M. G. & Takizawa, H. Inflamm-aging of hematopoiesis, hematopoietic stem cells, and the bone
marrow microenvironment. _Front. Immunol_. https://doi.org/10.3389/fimmu.2016.00502 (2016). * Soysal, P., Arik, F., Smith, L., Jackson, S. E. & Isik, A. T. in _Frailty and
Cardiovascular Diseases_ (ed. Veronese, N.) 55–64 (Springer International Publishing, 2020). * Almeida, M. & O’Brien, C. A. Basic biology of skeletal aging: role of stress response
pathways. _J. Gerontol. A. Biol. Sci. Med. Sci._ 68, 1197–1208 (2013). Article CAS Google Scholar * Ho, T. T. et al. Aged hematopoietic stem cells are refractory to bloodborne systemic
rejuvenation interventions. _J. Exp. Med._ 218, e20210223 (2021). Article CAS Google Scholar * Baryawno, N. et al. A cellular taxonomy of the bone marrow stroma in homeostasis and
leukemia. _Cell_ 177, 1915–1932.e16 (2019). Article CAS Google Scholar * Tikhonova, A. N. et al. The bone marrow microenvironment at single-cell resolution. _Nature_ 569, 222–228 (2019).
Article CAS Google Scholar * Baccin, C. et al. Combined single-cell and spatial transcriptomics reveal the molecular, cellular and spatial bone marrow niche organization. _Nat. Cell
Biol._ 22, 38–48 (2020). Article CAS Google Scholar * Kokkaliaris, K. D. et al. Adult blood stem cell localization reflects the abundance of reported bone marrow niche cell types and
their combinations. _Blood_ 136, 2296–2307 (2020). Article Google Scholar * Kusumbe, A. P. et al. Age-dependent modulation of vascular niches for haematopoietic stem cells. _Nature_ 532,
380–384 (2016). Article CAS Google Scholar * Poulos, M. G. et al. Endothelial transplantation rejuvenates aged hematopoietic stem cell function. _J. Clin. Invest._ 127, 4163–4178 (2017).
Article Google Scholar * Maryanovich, M. et al. Adrenergic nerve degeneration in bone marrow drives aging of the hematopoietic stem cell niche. _Nat. Med._ 24, 782–791 (2018). Article CAS
Google Scholar * Ho, Y.-H. et al. Remodeling of bone marrow hematopoietic stem cell niches promotes myeloid cell expansion during premature or physiological aging. _Cell Stem Cell_ 25,
407–418.e6 (2019). Article CAS Google Scholar * Guidi, N. et al. Osteopontin attenuates aging‐associated phenotypes of hematopoietic stem cells. _EMBO J._ 36, 840–853 (2017). Article CAS
Google Scholar * Shen, B. et al. A mechanosensitive peri-arteriolar niche for osteogenesis and lymphopoiesis. _Nature_ 591, 438–444 (2021). Article CAS Google Scholar * Ergen, A. V.,
Boles, N. C. & Goodell, M. A. Rantes/Ccl5 influences hematopoietic stem cell subtypes and causes myeloid skewing. _Blood_ 119, 2500–2509 (2012). Article CAS Google Scholar *
Kovtonyuk, L. V. et al. IL-1 mediates microbiome-induced inflammaging of hematopoietic stem cells in mice. _Blood_ 139, 44–58 (2022). Article CAS Google Scholar * Grover, A. et al.
Single-cell RNA sequencing reveals molecular and functional platelet bias of aged haematopoietic stem cells. _Nat. Commun._ 7, 11075 (2016). Article CAS Google Scholar * Poscablo, D. M.,
Worthington, A. K., Smith-Berdan, S. & Forsberg, E. C. Megakaryocyte progenitor cell function is enhanced upon aging despite the functional decline of aged hematopoietic stem cells.
_Stem Cell Rep._ 16, 1598–1613 (2021). Article CAS Google Scholar * Glatt, V., Canalis, E., Stadmeyer, L. & Bouxsein, M. L. Age-related changes in trabecular architecture differ in
female and male C57BL/6J Mice. _J. Bone Miner. Res._ 22, 1197–1207 (2007). Article Google Scholar * Schepers, K. et al. Myeloproliferative neoplasia remodels the endosteal bone marrow
niche into a self-reinforcing leukemic niche. _Cell Stem Cell_ 13, 285–299 (2013). Article CAS Google Scholar * Xu, X. et al. Age-related impairment of vascular structure and functions.
_Aging Dis._ 8, 590 (2017). Article Google Scholar * Ambrosi, T. H. et al. Aged skeletal stem cells generate an inflammatory degenerative niche. _Nature_ 597, 256–262 (2021). Article CAS
Google Scholar * Nishikawa, K. et al. Maf promotes osteoblast differentiation in mice by mediating the age-related switch in mesenchymal cell differentiation. _J. Clin. Invest._ 120,
3455–3465 (2010). Article CAS Google Scholar * Singh, L. et al. Aging alters bone-fat reciprocity by shifting in vivo mesenchymal precursor cell fate towards an adipogenic lineage. _Bone_
85, 29–36 (2016). Article CAS Google Scholar * Frisch, B. J. et al. Aged marrow macrophages expand platelet-biased hematopoietic stem cells via interleukin-1B. _JCI Insight_ 4, e124213
(2019). Article Google Scholar * Villeda, S. A. et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. _Nature_ 477, 90–94 (2011). Article CAS Google
Scholar * Mrak, R. Interleukin-1, neuroinflammation, and Alzheimer’s disease. _Neurobiol. Aging_ 22, 903–908 (2001). Article CAS Google Scholar * Woollard, K. J. Soluble bio-markers in
vascular disease: much more than gauges of disease? _Clin. Exp. Pharmacol. Physiol._ 32, 233–240 (2005). Article CAS Google Scholar * Abdelmagid, S. M. et al. Mutation in osteoactivin
decreases bone formation in vivo and osteoblast differentiation in vitro. _Am. J. Pathol._ 184, 697–713 (2014). Article CAS Google Scholar * Villarroya, F., Cereijo, R., Villarroya, J.
& Giralt, M. Brown adipose tissue as a secretory organ. _Nat. Rev. Endocrinol._ 13, 26–35 (2017). Article CAS Google Scholar * Yue, R., Zhou, B. O., Shimada, I. S., Zhao, Z. &
Morrison, S. J. Leptin receptor promotes adipogenesis and reduces osteogenesis by regulating mesenchymal stromal cells in adult bone marrow. _Cell Stem Cell_ 18, 782–796 (2016). Article CAS
Google Scholar * Coppé, J.-P., Desprez, P.-Y., Krtolica, A. & Campisi, J. The senescence-associated secretory phenotype: the dark side of tumor suppression. _Annu. Rev. Pathol. Mech.
Dis._ 5, 99–118 (2010). Article Google Scholar * Pietras, E. M. et al. Chronic interleukin-1 exposure drives haematopoietic stem cells towards precocious myeloid differentiation at the
expense of self-renewal. _Nat. Cell Biol._ 18, 607–618 (2016). Article CAS Google Scholar * Gekas, C. & Graf, T. CD41 expression marks myeloid-biased adult hematopoietic stem cells
and increases with age. _Blood_ 121, 4463–4472 (2013). Article CAS Google Scholar * Young, K. et al. Progressive alterations in multipotent hematopoietic progenitors underlie lymphoid
cell loss in aging. _J. Exp. Med._ 213, 2259–2267 (2016). Article CAS Google Scholar * Rodriguez-Fraticelli, A. E. et al. Clonal analysis of lineage fate in native haematopoiesis.
_Nature_ 553, 212–216 (2018). Article CAS Google Scholar * Dong, F. et al. Differentiation of transplanted haematopoietic stem cells tracked by single-cell transcriptomic analysis. _Nat.
Cell Biol._ 22, 630–639 (2020). Article CAS Google Scholar * Flach, J. et al. Replication stress is a potent driver of functional decline in ageing haematopoietic stem cells. _Nature_
512, 198–202 (2014). Article CAS Google Scholar * Gutierrez-Martinez, P. et al. Diminished apoptotic priming and ATM signalling confer a survival advantage onto aged haematopoietic stem
cells in response to DNA damage. _Nat. Cell Biol._ 20, 413–421 (2018). Article CAS Google Scholar * Hérault, A. et al. Myeloid progenitor cluster formation drives emergency and leukaemic
myelopoiesis. _Nature_ 544, 53–58 (2017). Article Google Scholar * Park, D. et al. Endogenous bone marrow MSCs are dynamic, fate-restricted participants in bone maintenance and
regeneration. _Cell Stem Cell_ 10, 259–272 (2012). Article CAS Google Scholar * Yamashita, M. & Passegué, E. TNF-α coordinates hematopoietic stem cell survival and myeloid
regeneration. _Cell Stem Cell_ 25, 357–372.e7 (2019). Article CAS Google Scholar * Furman, D. et al. Expression of specific inflammasome gene modules stratifies older individuals into two
extreme clinical and immunological states. _Nat. Med._ 23, 174–184 (2017). Article CAS Google Scholar * Zhou, B. O., Yue, R., Murphy, M. M., Peyer, J. G. & Morrison, S. J.
Leptin-receptor-expressing mesenchymal stromal cells represent the main source of bone formed by adult bone marrow. _Cell Stem Cell_ 15, 154–168 (2014). Article CAS Google Scholar *
Picelli, S. et al. Full-length RNA-seq from single cells using Smart-seq2. _Nat. Protoc._ 9, 171–181 (2014). Article CAS Google Scholar * Bagnoli, J. W. et al. Sensitive and powerful
single-cell RNA sequencing using mcSCRB-seq. _Nat. Commun._ 9, 2937 (2018). Article Google Scholar * Anders, S., Pyl, P. T. & Huber, W. HTSeq—a Python framework to work with
high-throughput sequencing data. _Bioinformatics_ 31, 166–169 (2015). Article CAS Google Scholar * Brennecke, P. et al. Accounting for technical noise in single-cell RNA-seq experiments.
_Nat. Methods_ 10, 1093–1095 (2013). Article CAS Google Scholar * Olsson, A. et al. Single-cell analysis of mixed-lineage states leading to a binary cell fate choice. _Nature_ 537,
698–702 (2016). Article CAS Google Scholar * Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. _Genome Biol._ 15,
550 (2014). Article Google Scholar * Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. _Genome Biol._ 19, 15 (2018). Article
Google Scholar * Wolock, S. L., Lopez, R. & Klein, A. M. Scrublet: computational identification of cell doublets in single-cell transcriptomic data. _Cell Syst._ 8, 281–291.e9 (2019).
Article CAS Google Scholar * Hoffman, P. et al. Comprehensive Integration of Single-Cell Data. _Cell_ 177, 1888–1902 (2019). Article Google Scholar * Hao, Y. et al. Integrated analysis
of multimodal single-cell data. _Cell_ 184, 3573–3587.e29 (2021). Article CAS Google Scholar * Tusher, V. G., Tibshirani, R. & Chu, G. Significance analysis of microarrays applied to
the ionizing radiation response. _Proc. Natl Acad. Sci. USA_ 98, 5116–5121 (2001). Article CAS Google Scholar Download references ACKNOWLEDGEMENTS We thank S. Villeda (UCSF) for providing
some of the old C57BL/6 mice; D. Reynaud (UCSF) for initial analyses of the old mice; A. Valencia (UCSF) for technical assistance with various experiments; S. Kinston (Cambridge University)
for scRNA-seq library preparations; A. Li (Bone Imaging Research Core, UCSF) and both E. Guo and P. T. Shyu (CUIMC) for microCT analyses; M. Lee (UCSF) and M. Kissner (CUIMC) for management
of our Flow Cytometry Core facilities; and all members of the Passegué Laboratory for critical insights and suggestions. C.A.M. was supported by NIH F31HL160207 and a NYSTEM training grant;
E.V.V. by a Rubicon Grant from The Netherlands Organization for Scientific Research, a Stem Cell Grant from BD Biosciences and a NYSTEM training grant; O.C.O by CRI/Margaret Dammann Eisner
postdoctoral fellowship CRI3617; J.W.S. by long-term EMBO postdoctoral fellowship ALTF-2021-196; P.V.D. by NIH F31HL151140; E.M.P. by NIH F32HL106989 and K01DK09831; S.T.B. by a CIRM
postdoctoral fellowship; and T.T.H. by an AHA and Hillblom Center for the Biology of Aging predoctoral fellowship. F.J.C-N., X.W. and B.G. were supported by grants from the Wellcome
(206328/Z/17/Z), CRUK (C1163/A21762) and core funding by the Wellcome to the Cambridge Stem Cell Institute. This work was funded by NIH R01CA184014, NIH R35HL135763, Glenn Foundation
Research Award and LLS Scholar Award to E.P., and supported in part through the NIH/NCI Cancer Center Support Grant P30CA013696 to CUIMC. The funders of this work had no role in study
design, data collection and analysis, decision to publish or preparation of the manuscript. Clipart in figures was created using BioRender. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS *
Columbia Stem Cell Initiative, Department of Genetics and Development, Columbia University Irving Medical Center, New York, NY, USA Carl A. Mitchell, Evgenia V. Verovskaya, Oakley C. Olson,
James W. Swann, Paul V. Dellorusso & Emmanuelle Passegué * The Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell Research, Department of Medicine, Division
Hematology/Oncology, University of California San Francisco, San Francisco, CA, USA Evgenia V. Verovskaya, Aurélie Hérault, Si Yi Zhang, Arthur Flohr Svendsen, Eric M. Pietras, Sietske T.
Bakker, Theodore T. Ho & Emmanuelle Passegué * Wellcome and MRC Cambridge Stem Cell Institute, Department of Haematology, Jeffrey Cheah Biomedical Centre, Cambridge University,
Cambridge, UK Fernando J. Calero-Nieto, Xiaonan Wang & Berthold Göttgens Authors * Carl A. Mitchell View author publications You can also search for this author inPubMed Google Scholar *
Evgenia V. Verovskaya View author publications You can also search for this author inPubMed Google Scholar * Fernando J. Calero-Nieto View author publications You can also search for this
author inPubMed Google Scholar * Oakley C. Olson View author publications You can also search for this author inPubMed Google Scholar * James W. Swann View author publications You can also
search for this author inPubMed Google Scholar * Xiaonan Wang View author publications You can also search for this author inPubMed Google Scholar * Aurélie Hérault View author publications
You can also search for this author inPubMed Google Scholar * Paul V. Dellorusso View author publications You can also search for this author inPubMed Google Scholar * Si Yi Zhang View
author publications You can also search for this author inPubMed Google Scholar * Arthur Flohr Svendsen View author publications You can also search for this author inPubMed Google Scholar *
Eric M. Pietras View author publications You can also search for this author inPubMed Google Scholar * Sietske T. Bakker View author publications You can also search for this author
inPubMed Google Scholar * Theodore T. Ho View author publications You can also search for this author inPubMed Google Scholar * Berthold Göttgens View author publications You can also search
for this author inPubMed Google Scholar * Emmanuelle Passegué View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS E.V.V. initiated the studies
and contributed to most of the experiments with help from A.H. for BM fluid isolation and immunophenotyping. S.Y.Z. performed GMP cluster analyses and ELISA assays. A.F.S. performed
immunofluorescence and whole-mount staining. S.T.B. performed initial 5-FU analyses. T.T.H. performed cell collection and provided technical assistance. C.A.M. re-analysed all the generated
data, contributed to various stromal and molecular analyses and, with help from O.C.O., J.W.S. and P.V.D, completed the chronic IL-1β exposure studies and performed all the analyses of
_Il1r1_-deficient mice. F.J.C-N., X.W. and B.G. prepared and analysed the scRNA-seq samples. E.M.P. prepared the microarray samples. E.V.V. and E.P. designed the initial experiments, and
C.A.M. and E.P. revised the experiments. E.V.V., C.A.M. and E.P. wrote and edited the manuscript. C.A.M. and E.P. handled the resubmission. CORRESPONDING AUTHOR Correspondence to Emmanuelle
Passegué. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Cell Biology_ thanks Iannis Aifantis and the other,
anonymous, reviewer(s) for their contribution to the peer review of this work. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in
published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 GROSS ANALYSIS OF THE REMODELED OLD BM CAVITY. A, H&E staining of humeri and sterna of 2 individual
young and old mice. Scale bar, 100 µm. B, TPO and TGF-β levels in young and old BM fluids. Results are from 2 independent cohorts. C, μCT analyses of young and old tibia with representative
images of cortical and trabecular regions (left) and quantification of bone volume/total volume (BV/TV) and connectivity density (right). D, Representative image of bone lining ALCAM+
osteoblasts immunofluorescence staining in young and old mice. Representative of 3 independent experiments. Scale bar, 100 µm. E, Representative images and quantification of
immunofluorescence staining of vascular volume (laminin) and vascular leakage by dragon-green beads (DGB) diffusion assay in young and old BM. Scale bar, 50 µm. F, Representative images and
quantification by flow cytometry of DGB endocytosis in young and old marrow SEC; scale bar, 5 µm. Results are from 3 independent cohorts. G, Von Kossa staining in young and old endosteal
MSC-S. H, Experimental scheme for the indicated co-culture experiments showing the effects of young or old BM cells on young MSC-S (top right), and young BM cells on young and old MSC-S
(bottom). I, Frequency of endosteal and marrow mesenchymal populations in young and middle age (13-month-old) mice with changes in CFU-F from endosteal OLCs (bottom right). Data are means ±
S.D; P-values were obtained by two-tailed Welch’s t-test without adjustment for multiple comparisons. Source data EXTENDED DATA FIG. 2 MOLECULAR FEATURES OF OLD MESENCHYMAL POPULATIONS. A,
UMAP visualization of the entire plate-based scRNAseq dataset of niche populations of mesenchymal and endothelial populations shown in Fig. 2a. B, Global changes in MSC-L gene identity in
clusters M2 vs. M1. C, Representative flow cytometry staining (top) and quantification of LepR and PDGF-Rα levels (bottom) in young and old MSC-L. Results are from 6 independently analyzed
groups of 1 or 2 young or old mice. D, HSC niche factors with violin plots representation of _Kitl_ and _Cxcl12_ expression in the indicated young and old mesenchymal populations (left) and
SCF and SDF1α levels in young and old BM fluids (right). E, GSEA results for Hallmark biological processes significantly affected in old MSC-L-like, OPr-like and MSC-S-like groups (FDR <
0.05). Data are means ± S.D. except for (b) and (d); P-values were obtained by two-tailed Student’s t-test without adjustment for multiple comparisons (b), or by Welch’s t-test without
adjustment for multiple comparisons (c,d). Source data EXTENDED DATA FIG. 3 MOLECULAR FEATURES OF OLD ENDOTHELIAL POPULATIONS AND FURTHER CHARACTERISTICS OF THE OLD NICHE. A, ICGS output of
young and old endothelial populations from the plate-based scRNAseq dataset shown in Fig. 2a with 7 clusters of cells (E1 to E7, horizontal) defined according to the expressing pattern of
the 7 clusters of genes (a to g, vertical). Examples included in gene clusters a to g are shown. Star, contaminating mesenchymal/endothelial doublets; P, pericytes; A, arterioles; T,
transition vessels.Color scheme is based on normalized gene expression level from the indicated minimum(blue) to maximum (yellow) value in the scale. B, Ingenuity Pathway Analysis (IPA)
canonical pathways enriched in old AEC-like and SEC-like groups (Z-score > 1; p < 0.01). C, Characteristic expression patterns for the indicated genes in the droplet-based scRNAseq
dataset of young and old endosteal and central marrow stromal fractions shown in Fig. 2c. Cells in the UMAP were colored according to the expression levels of the indicated genes. Color
scheme is based on ln scale of normalized counts from 0 (gray) to the indicated maximum value in the scale (red). Source data EXTENDED DATA FIG. 4 AGE-RELATED CHANGES IN BLOOD AND BM
POPULATIONS AND ALTERED LINEAGE BIAS IN OLD MPPS. A, Overlap between cytokines upregulated in old BM fluids and published SASP profile40. B, Representative SA-β-gal staining in control
irradiated mouse embryonic fibroblasts (MEF) and isolated young and old MSC-S and OLC. Representative of 3 independent experiments. Scale bar, 20 µm. C, Absolute expression of _Il1b_ in
different mature hematopoietic cell types and unfractionated endosteal and central marrow stroma from pooled young and old samples. Results are expressed as -log(dCt) relative to _gapdh_. D,
Complete blood count (CBC) parameters in young and old mice. Results are from 7 independent cohorts. WBC, white blood cell; My, myeloid; Ly, lymphocyte; RBC, red blood cell; Pt, platelet.
E, Cellularity and quantification of mature populations in young and old BM. Results are from 3 independent cohorts. Gr, granulocyte. F, Quantification of progenitor populations in young and
old BM. Results are from 3 independent cohorts. CMP, common myeloid progenitor; GMP, granulocyte-macrophage progenitor; MEP, megakaryocyte-erythrocyte progenitor; CLP, common lymphoid
progenitor; CFU-E, erythroid colony-forming unit; Pre-GM, pre-granulocyte/macrophage; Pre-MegE, pre-megakaryocyte/erythrocyte; MkP, megakaryocyte progenitor. G, Representative staining and
quantification of young and old HSC markers. Results are from 2 independent cohorts, with data represented as box and whiskers (min to max) and expressed as fold changes in mean fluorescence
intensity (MFI) relative to young HSCs. H, Characteristic expression patterns of lineage determinant genes in the droplet-based scRNAseq young and old LK/LSK dataset. Cells in the UMAP were
colored according to the expression levels of the indicated genes. I, IPA Upstream Regulators analysis of young and old populations from the droplet-based scRNAseq young and old LK/LSK
dataset filtered on cytokines and growth factors. Data are means ± S.D. except for (g) where box ranges from 25th to 75th percentile with center line at the median, and whiskers range from
minima to maxima; P-values were obtained by two-tailed Welch’s t-test without adjustment for multiple comparisons. Source data EXTENDED DATA FIG. 5 ALTERED FUNCTIONALITY OF OLD HSCS AND
MPPS. A, Colony formation in methylcellulose for young (Y) and old (O) HSCs, MPP3 and MPP4. Results are from 2 independent experiments. Mix: all lineages; GM: granulocyte/macrophage; G(or)M:
granulocyte (or) macrophage; MegE: megakaryocyte/erythrocyte; CFU: colony-forming units. B, Myeloid differentiation in liquid culture for young and old MPP3 with quantification (right) of
immature Sca-1+/c-Kit+ cells (left) and mature Mac-1+/FcγR+ macrophage (right). C, Representative flow cytometry staining of CD19+ lymphoid vs. Mac-1+ myeloid differentiation in OP9 + IL7
culture conditions for young and old HSCs, MPP3 and MPP4. Results are representative of 3 independent experiments. D, Representative histograms of CFSE staining of cultured young and old
HSCs, MPP3 and MPP4. Results are representative of 3 independent experiments. E, Cleaved caspase 3/7 (CC3/7) activity in cultured young and old HSCs, MPP3 and MPP4. F, Short-term lineage
tracking following transplantations of young and old HSCs, MPP3 and MPP4 in sub-lethally irradiated recipients with experimental scheme (left) and quantification of overall blood donor
chimerism (top graphs) and myeloid chimerism among donor cells (bottom graphs). Results are from 3 independent cohorts. Data are means ± S.D. except for (f) (± S.E.M.); P-values were
obtained by two-tailed Welch’s t-test without adjustment for multiple comparisons (a, d, f), or by two-tailed Student’s t-test without adjustment for multiple comparisons (b). Source data
EXTENDED DATA FIG. 6 IMPROVED AGING FEATURES WITH IL-1 SIGNALING BLOCKADE. A–C, Short-term blockade of IL-1 signaling upon Anakinra (Ana) treatment in young (Y) and old (O) mice with: (a)
experimental scheme; (b) changes in HPSC frequency; and (c) engraftment over time (left) and lineage reconstitution (right) at 4 months (4 mo) post-transplantation (Tplx) of the indicated
HSC populations. Results are from 3 independent cohorts of young and old mice injected with either PBS or Anakinra, with HSCs isolated from the pooled BM of mice from the same treatment
group and transplanted into 3 to 5 recipients, each. D, E, Additional characterization of the effects of Anakinra blockade of IL-1 signaling during 5FU-mediated regeneration in young and old
mice with: (d) changes in IL-1α, IL-1β and MIP1α levels in BM fluids; and (e) platelet (Plt) levels in peripheral blood. Results are from 3 independent cohorts started with 15 young and 11
old mice treated once with 5FU, injected daily with either PBS or Anakinra, and analyzed at day 12 post-5FU treatment. F, GSEA results for Hallmark biological processes significantly
enriched in MSC-L2 vs. MSC-L1 groups (FDR < 0.05). Data are from the droplet-based scRNAseq analyses of endosteal and central marrow stromal fractions in young (n = 2) and old (n = 2)
_Il1r1__+/+_ wild type (WT) mice and old (n = 1) _Il1r1__−/−_ mice shown in Fig. 7b. G, H, Droplet-based scRNAseq analyses of Lin−/c-Kit+ (LK) and Lin−/Sca-1+/c-Kit+ (LSK) BM fractions
isolated from young (n = 2) and old (n = 2) _Il1r1__+/+_ wild type (WT) and old (n = 1) _Il1r1__−/−_ mice with (g) UMAP visualization and (h) quantification of percent of HSPCs and
progenitors. Data are means ± S.D. except for engraftment results shown in (c) (± S.E.M.); P-values were obtained by one-way Anova adjusted for multiple comparisons using the Holm-Šídák
method. Source data EXTENDED DATA FIG. 7 KEY ROLE OF IL-1 IN THE AGING OF BOTH BM NICHE AND BLOOD SYSTEM. A, B, GSEA results in old _Il1r1__−/−_ HSPC population for the Gene Ontology
pathways affected in either old WT HSPCs (a) or young WT HSPCs (b) identified by droplet-based scRNAseq analyses. n/a, non-available; ns, not significant; *nominal p value ≤ 0.05, ** nominal
p value ≤ 0.01, *** nominal p value ≤ 0.001. C, Peripheral blood CD45.1+ donor chimerism (left) and number of donor-derived GMPs (right) in young or old WT and _Il1r1__−/−_ CD45.2+
recipient mice at 4 months (mo) after lethal irradiation and transplantation (Tplx) with 2 × 106 young WT CD45.1+ donor BM cells. D, _Il1r1_ expression in the droplet-based scRNAseq of young
and old WT stroma and LK/LSK datasets. Cells in the UMAP were colored according to the expression levels of the indicated genes. Color scheme is based on ln scale of normalized counts from
the indicated minimum (gray) to maximum (red) value in the scale. E–I, Unchanged aging features in old _Tnf__−/−_ mice with: (e) color scheme; (f) blood parameters; (g) endosteal (left) and
central marrow (right) mesenchymal population frequencies; (h) BM hematopoietic population frequencies; and (i) engraftment over time (left) and lineage reconstitution (right) at 4 mo
post-Tplx of the indicated HSC populations. Results are from 3 independent cohorts of young and old WT and age-matched _Tnf__−/−_ mice, with HSCs isolated from the pooled BM of mice of the
same genotype and transplanted into 3 to 5 recipients, each. Data are means ± S.D. except for engraftment results shown in (i) (± S.E.M.); P-values were obtained by Kolmogorov-Smirnov test
(a,b), by one-way Anova adjusted for multiple comparisons using the Holm-Šídák method (c,i), or by two-tailed Student’s t-test without adjustment for multiple comparisons (f, g, h). Source
data EXTENDED DATA FIG. 8 SCHEMATIC OF THE CROSSTALK BETWEEN THE BM NICHE AND HEMATOPOIETIC SYSTEM DURING PHYSIOLOGICAL AGING. In youth, HSCs reside primarily in the central marrow where
they are maintained by peri-sinusoidal MSC-L and produce a balanced output of all mature cell lineages (Mk, megakaryocytes; Ery, erythrocytes, My, myeloid cells; Ly, lymphoid cells).
Abundant peri-arteriolar MSC-S at the endosteum efficiently produce OPr cells that support osteoblast development, ECM deposition and bone formation. With age, numerical loss and functional
decline of MSC-S and OPr leads to bone thinning, with the remaining OPr constitutively producing IL-1. Chronic IL-1, in turn, reinforces niche degradation at the endosteum and contributes to
dysfunction of the sinusoidal vasculature. Chronic IL-1 also acts in trans on central marrow MSC-L and HSPCs, driving the appearance of an inflammatory iMSC-L subset and steady-state
engagement of emergency myelopoiesis (EM) programs with GMP cluster (cGMP) formation. Strikingly, acute IL-1 blockade with Anakinra enables more youthful blood production during 5FU-mediated
regeneration, and life-long removal of IL-1 signaling in _Il1r1__−/−_ mice maintains MSC-L in a more youthful cell state associated with improved blood production and HSC function.
SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURES Supplementary Figs. 1–6 REPORTING SUMMARY SUPPLEMENTARY TABLES Supplementary Table 1: ICGS of young and old niche cells. Plate-based scRNA-seq
gene expression data of mesenchymal and ECs were separated and subjected to unsupervised single-cell population identification using ICGS. The relative expression of every guide gene in
each cluster, as well as the relative expression of every guide gene in each pooled group of clusters corresponding to the indicated cell types, was calculated by averaging the relative
expression of each gene across individual cells within a cluster or group. Supplementary Table 2: DEGs and pathway analyses of young versus old niche cells. Plate-based scRNA-seq gene
expression data between young and old cells in pooled identity groups as defined by ICGS were analysed for DEGs using the DESeq2 package. Overlap with Hallmark gene sets was evaluated, and
significant (FDR < 0.05) overlaps are indicated in bold for mesenchymal populations. As no significant overlaps were observed for endothelial populations, IPA software (Qiagen) was used
to determine IPA canonical pathways with absolute _z_-score ≥ 1 and –log10(_P_ value) ≥ 2 enriched in young and old AEC-like and SEC-like cells. Supplementary Table 3: Quantibody array-based
measurements of cytokine concentration in young and old BM fluids. BM fluids isolated from young (10 weeks of age) and old (27–29 months of age) mice (_n_ = 5) were analysed using
Quantibody Testing Service (Raybiotech). Samples were diluted fourfold before analyses. Results are mean ± s.d. and are expressed as pg ml–1 concentration; *_P_ < 0.05, **_P_ _<_ 0.05,
***_P_ < 0.05. Supplementary Table 4: DEGs and pathway analyses of young versus old HSCs. SAM was performed on young and old cells within each population to determine SAM delta scores.
The top 1,000 most highly DEGs for each group were collected. SAM scores for these genes were used for GSEA and overlap with Reactome gene sets was evaluated for each population. The top 5
statistically significant (FDR < 0.05) overlaps for each group are in bold. SOURCE DATA SOURCE DATA FIG. 1 Statistical source data. SOURCE DATA FIG. 3 Statistical source data. SOURCE DATA
FIG. 4 Statistical source data. SOURCE DATA FIG. 5 Statistical source data. SOURCE DATA FIG. 6 Statistical source data. SOURCE DATA FIG. 7 Statistical source data. SOURCE DATA EXTENDED DATA
FIG. 1 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 2 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 3 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 4
Statistical source data. SOURCE DATA EXTENDED DATA FIG. 5 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 6 Statistical source data. SOURCE DATA EXTENDED DATA FIG. 7 Statistical
source data. RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Mitchell, C.A., Verovskaya, E.V., Calero-Nieto, F.J. _et al._ Stromal niche inflammation
mediated by IL-1 signalling is a targetable driver of haematopoietic ageing. _Nat Cell Biol_ 25, 30–41 (2023). https://doi.org/10.1038/s41556-022-01053-0 Download citation * Received: 09
November 2021 * Accepted: 15 November 2022 * Published: 17 January 2023 * Issue Date: January 2023 * DOI: https://doi.org/10.1038/s41556-022-01053-0 SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative