Play all audios:
ABSTRACT The rice blast fungus _Magnaporthe oryzae_ uses a pressurized infection cell called an appressorium to drive a rigid penetration peg through the leaf cuticle. The vast internal
pressure of an appressorium is very challenging to investigate, leaving our understanding of the cellular mechanics of plant infection incomplete. Here, using fluorescence lifetime imaging
of a membrane-targeting molecular mechanoprobe, we quantify changes in membrane tension in _M. oryzae_. We show that extreme pressure in the appressorium leads to large-scale spatial
heterogeneities in membrane mechanics, much greater than those observed in any cell type previously. By contrast, non-pathogenic melanin-deficient mutants, exhibit low spatially homogeneous
membrane tension. The sensor kinase ∆_sln1_ mutant displays significantly higher membrane tension during inflation of the appressorium, providing evidence that Sln1 controls turgor
throughout plant infection. This non-invasive, live cell imaging technique therefore provides new insight into the enormous invasive forces deployed by pathogenic fungi to invade their
hosts, offering the potential for new disease intervention strategies. SIMILAR CONTENT BEING VIEWED BY OTHERS REVEALING REAL-TIME 3D IN VIVO PATHOGEN DYNAMICS IN PLANTS BY LABEL-FREE OPTICAL
COHERENCE TOMOGRAPHY Article Open access 27 September 2024 DISSECTING THE MEMBRANE-MICROTUBULE SENSOR IN GRAPEVINE DEFENCE Article Open access 01 December 2021 CARBOTAG: A MODULAR APPROACH
FOR LIVE AND FUNCTIONAL IMAGING OF PLANT CELL WALLS Article Open access 01 May 2025 MAIN Many plant pathogens use specialized infection cells called appressoria to infect their hosts1,2,3.
Appressoria facilitate pathogen entry into host tissue to cause disease, and the famous ‘gold leaf’ experiment demonstrated the capacity of some fungal appressoria to puncture the leaf
surface using force generation rather than enzymatic activity2,4. Appressoria of the rice blast fungus _Magnaporthe oryzae_ (synonym of _Pyricularia oryzae_)5—a major threat to global food
security6,7,8—breach the tough surface of rice leaves and, remarkably, other hard synthetic surfaces by generating turgor of up to 8.0 MPa (~40 times the pressure of a car tyre9). This
generates force at the base of the appressorium, measured in a related pathogen _Colletotrichum graminicola_ using an optical waveguide, of 17 µN (ref. 10). By contrast, pressures measured
in fungal or oomycete hyphae seldom exceed 0.8 MPa (ref. 11). _M. oryzae_ appressoria have a melanin-lined cell wall impermeable to glycerol, but freely permeable to water, which rapidly
enters the cell, generating hydrostatic pressure. Mutation of the melanin biosynthetic enzyme-encoding genes _ALB1, RSY1_ and _BUF1_ causes loss of appressorium melanization, which enables
movement of solutes and water through the appressorium cell wall, leading to loss of turgor generation and the ability to cause disease12,13. Directly measuring appressorium turgor has been
challenging because of the enormous pressure generated, which preclude the use of pressure probes. Instead, researchers have used indirect methods such as the incipient cytorrhysis assay,
which records the rate of cell collapse when appressoria are incubated in hyperosmotic solutions9,14,15. However, melanin-deficient mutants undergo plasmolysis rather than cell collapse when
exposed to high concentrations of glycerol limiting the use of this assay9,14. The Flipper-TR probe, which contains a membrane-targeted twisted push–pull fluorophore is sensitive to
mechanical forces acting on the plasma membrane. Previous reports have suggested the fluorescence lifetime of the probe changes linearly with plasma membrane tension in both yeast and
mammalian cells16. In _M. oryzae_, the probe has been used to measure plasma membrane tension in hyphae of Guy11 and a _∆vast1_ mutant, which affects TOR17 signalling, and is implicated in
the cAMP response, cell integrity and control of autophagy18,19,20,21,22. But, while the probe suggests that the _∆vast1_ mutant has increased tension, experiments were performed only in
hyphae23. Here we set out to explore whether we could visualize turgor generation in appressoria directly. Recently, a set of chemically modified molecular rotors were developed to yield
complete microviscosity maps of cells and tissues in the cytosol, vacuole, plasma membrane and wall of plant cells24. These boron-dipyrromethene-based molecular rotors are rigidochromic,
meaning their fluorescence lifetime depends on the mechanics of their surroundings, such as viscosity or membrane tension. The N+-BDP plasma membrane probe has, for instance, revealed
differences in membrane mechanics between the plant root cap and the meristem. Fluorescence lifetime imaging microscopy (FLIM) revealed the plant meristem to undergo continuous growth and
cell division, resulting in constant tension in the plasma membrane24. The tension increases spacing between lipids, leading to significant reduction in membrane rotor lifetime compared to
the relaxed plasma membranes of root cap cells25. Furthermore, closer examination of the plasma membrane revealed distinct lipid microdomains within a single bilayer. Likewise, in root hairs
the fluorescence lifetime was lower at the growing tip (3.6 ± 0.8 ns), compared with the non-growing epidermal cell plasma membrane (4.3 ± 0.6 ns). The change in lifetime corresponds to the
increase in tension in the growing root hair tip, where membrane curvature is greatest. Plasmolysis assays in rotor-stained root hairs confirmed the probe’s responsiveness to changes in
tension within _Arabidopsis_ root tissues, as the fluorescence lifetime within root hair tips significantly increased upon exposure to hyperosmotic stress and a drop in membrane tension. In
this Article, we report that the mechanosensor N+-BDP plasma membrane rotor probe can detect spatial variations in membrane tension in _M. oryzae_ appressoria. We reveal how changes in
microviscosity correlate with appressorium-mediated turgor-driven plant cell infection and show that, under the extreme pressures of an appressorium, the plasma membrane exhibits spatial
heterogeneity in tension, a phenomenon not previously observed in living cells. We provide experimental validation of the requirement of melanin biosynthesis for appressorium turgor
generation in _M. oryzae_ and show that the Sln1 turgor sensor kinase26,27 is necessary for controlling both the rate of turgor generation and its modulation before plant infection. The
N+-BDP mechanosensor therefore provides a direct, quantitative measurement of the average tension at the appressorium membrane enabling new insight into the behaviour of eukaryotic cells
under extreme pressure and the mechanobiology of plant infection by the blast fungus. RESULTS N+-BDP REVEALS SPATIAL VARIATIONS IN PLASMA MEMBRANE TENSION IN _M. ORYZAE_ We first determined
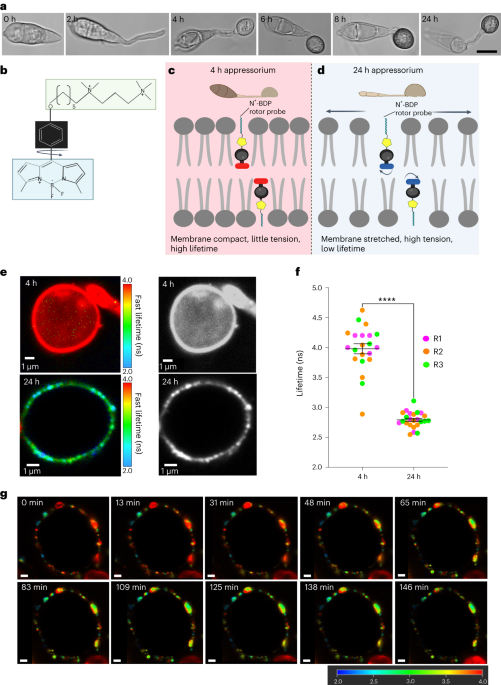
whether the mechanoprobe N+-BDP could reveal changes in appressorium-specific membrane tension during a time course of infection-related development of the wild-type _M. oryzae_ strain
Guy11. During initial stages of appressorium development, 4 h after conidia have germinated on hydrophobic glass coverslips, incipient appressoria are not melanized and have not yet
generated turgor. By contrast, 24 h after inoculation, appressoria are mature and fully melanized, and generate high levels of turgor, deployed as mechanical force as appressoria are tightly
bound to the hydrophobic surface, creating a tight seal necessary for appressorium function8,28,29 (Fig. 1a). The chemical structure of the mechanoprobe N+-BDP is based on a modified
phenyl-substituted boron-dipyrromethene molecular rotor, in which the phenyl ring carries an aliphatic tail with two permanent cationic charges, creating a positive charge and thereby
targeting the negatively charged phospholipid bilayer24 (Fig. 1b). Upon staining, the probe is positioned between the tails of the bilayer, with its aliphatic tail facing towards
phospholipid heads (Fig. 1c). Previous work using rotor-stained giant unilamellar vesicles (GUVs) composed of sphingomyelin, 1,2-dioleoyl-_sn_-glycero-3-phosphocholine and cholesterol
(0.56:0.24:0.20) has enabled study of lipid phase transition. The lipid phase separation in GUVs creates an inhomogeneous biological membrane composed of different lipid microdomains,
similar to formation of lipid microdomains in biological membranes by immiscibility of different lipids24,30,31,32. Upon staining different GUVs, the N+-BDP mechanoprobe demonstrated a
stronger mechanical restriction for rotations imposed by the tightly packed and solid-like sphingomyelin-rich gel-like ordered phase, generating longer average fluorescence lifetimes
compared with the less tightly packed liquid-like phase enriched in 1,2-dioleoyl-_sn_-glycero-3-phosphocholine. When considering an appressorium, we hypothesized that early-stage (4 h)
appressoria would display a more compact membrane as a result of being under little or no tension, thereby causing mechanical restriction of the rotor probe upon photoexcitation and longer
average fluorescence lifetimes (Fig. 1c). In a 24 h appressorium meanwhile exhibiting high appressorium turgor and high membrane tension, the membrane would become stretched and disordered,
allowing free rotation of the probe and consequently shorter average fluorescence lifetimes (Fig. 1d). To test this hypothesis, we used the N+-BDP mechanoprobe to stain 4 h and 24 h
appressoria of _M. oryzae_ Guy11 to observe the spatial variations in membrane tension (Fig. 1e). Strikingly, when we selected the appressorium membrane for analysis (Extended Data Fig. 1)
we observed that 4 h appressoria displayed a homogeneous high average rotor lifetime of 3.98 ± 0.084 ns (Fig. 1f and Supplementary Video 1), in contrast to mature appressoria (7.5–24 h)
which displayed spatial heterogeneity and a significantly lower average rotor lifetime of 2.79 ± 0.026 ns (Fig. 1f and Supplementary Video 2). Intriguingly, in contrast to the low
fluorescence lifetimes we consistently observed in appressoria, we observed consistent and uniform high fluorescence lifetimes in _M. oryzae_ germ tubes (Extended Data Fig. 2). Considering
the primary function of the germ tube is to deliver the contents of the conidium to the developing appressorium for maturation, these cells have no requirement for extreme turgor generation,
which is corroborated by the rotor probe. We next tested whether artificially lowering the turgor of appressoria by incubation in hyperosmotic concentrations of glycerol, would
independently corroborate probe responsiveness to changes in membrane tension within an appressorium. Under hyperosmotic conditions, the fluorescence lifetime in a 24 h appressorium
significantly increased from 2.79 ± 0.041 ns to 3.10 ± 0.067 ns upon addition of exogenous 1 M glycerol (Extended Data Fig. 3). This change is consistent with a drop in tension as water
exits the appressorium by osmosis13,26. Considering melanin biosynthesis and deposition occur between 4 h and 8 h post-inoculation (hpi) on a glass coverslip8, we reasoned that this would
provide a suitable time to capture changes in local membrane tension and turgor. A real-time movie of a Guy11 appressorium stained with the N+-BDP rotor probe was captured during a 3 h
period, in which spatial variations in membrane tension were apparent and the overall fluorescence lifetime decreased (Fig. 1g, Supplementary Video 3 and Extended Data Fig. 4). Our initial
analyses of turgor generation were carried out on unyielding glass coverslips, so we were interested in observing appressorium turgor on surfaces that can be penetrated. As the N+-BDP
mechanoprobe stains all membranes it encounters, imaging appressoria on a leaf surface is technically challenging. We therefore inoculated sterile onion epidermis, pre-treated with
chloroform before washing in sterile water (Extended Data Fig. 5a), which can be readily penetrated by the fungus. We found that appressorial membranes displayed consistent spatial
heterogeneity (Extended Data Fig. 5b) and a very similar lifetime compared with those on an unyielding coverslip surface at 2.69 ± 0.083 ns (Extended Data Fig. 5c). We conclude that the
dynamics of appressorium turgor generation are similar on both yielding and unyielding surfaces. To determine whether the N+-BDP mechanoprobe can be applied to other appressorium-forming
fungal pathogens, we tested appressoria from the hemibiotrophic ascomycete fungus _Colletotrichum higginsianum_, which causes anthracnose disease on _Arabidopsis_33. During the initial
stages of appressorium development, once conidia have germinated on hydrophobic glass coverslips, incipient appressoria of _C. higginsianum_ are not melanized and have not yet generated
turgor. By contrast, 24 h after inoculation, appressoria are mature and fully melanized (Fig. 2a). We used the N+-BDP mechanoprobe to stain early-stage (non-melanized) (Fig. 2b) and
late-stage (melanized) (Fig. 2c) appressoria of _C. higginsianum_, to observe spatial variations in membrane tension. Early-stage appressoria displayed homogeneous high average rotor
lifetime of 4.301 ± 0.043 ns (Fig. 2d), in contrast to late-stage appressoria which displayed a significantly lower average rotor lifetime of 3.72 ± 0.085 ns, consistent with our findings
for _M. oryzae_. We conclude that the N+-BDP mechanoprobe can be widely deployed for generating membrane tension maps in appressorium-forming pathogens. To confirm that the surprising large
inhomogeneties observed in _M. oryzae_ 24 h appressoria are due to changes in membrane tension and not the result of formation of compositional membrane microdomains or dye interaction with
the hydrophobic melanin layer, we performed control experiments using a plasma membrane sensor NR12S, a solvatochromic Nile red-based probe34. In the same way as the N+-BDP probe, the Nile
red chromophore is functionalized with a lipid-like alkyl tail for membrane targeting (Extended Data Fig. 6a). This probe, however, is not sensitive to membrane tension, but rather exhibits
a shift in emission wavelength in response to changes in local chemical polarity, such as lipid and sterol composition of the membrane. Ratiometric imaging, in which the total emission of
the dye is split into two channels, provides a non-lifetime-based readout for the probe. Changes in membrane chemical composition and lipid phase affect the chemical polarity of the probe
microenvironment, initiating a change in the intensity ratio between the blue and red channels35 (Extended Data Fig. 6b,c). Previously, this probe was used for mapping spatial variation in
plasma membrane chemical polarity of _Phytophthora infestans_ germlings35. We observed that chemical polarity of appressoria becomes globally lower during appressorium maturation. In early 4
h appressoria, ratiometric imaging of the appressoria plasma membrane appears blue, indicating the membrane has a high chemical polarity that is is less ordered. (Extended Data Fig. 6d–j).
However, in 24 h appressoria, the plasma membrane appears yellow and red, indicating the membrane has a low polarity indicative of a higher lipid packing order. (Extended Data Fig. 6k–p).
Importantly, the magnified sections of the plasma membrane for both 4 h appressoria (Extended Data Fig. 6e,f,h,i) and 24 h appressoria (Extended Data Fig. 6l,m,o,p) shows the probe NR12S
displaying either homogeneous polarity, or polarity variations whose pattern is not consistent with the larger changes in tension we observe with the 4 h and 24 h appressoria stained with
the N+-BDP rotor (for comparison, see Extended Data Fig. 6). We conclude that N+-BDP reveals spatial changes in appressorium-specific tension during infection-related development of _M.
oryzae_ and that the observed patterns are not caused by concurrent inhomogeneities in membrane composition. MELANIN IS CRITICAL FOR _M. ORYZAE_ APPRESSORIUM TURGOR The synthesis of
dihydroxy-naphthalene (DHN) melanin in _M. oryzae_ has long been reported to be essential for appressorium-specific turgor-driven plant infection7,13. A layer of melanin is located between
the appressorium membrane and cell wall where it acts as a structural barrier to efflux of solutes from the appressorium, essential for turgor generation and pathogenicity7,13. Mutation of
genes encoding enzymes required for DHN-melanin biosynthesis, _ALB1_, _RSY1_ and _BUF1_, causes impairment in appressorium and hyphal melanization12. Consequently, melanin-deficient mutants
fail to infect intact host plants12. We determined whether N+-BDP could detect a reduction in membrane tension in _alb1__-_ and _buf1__-_ mutants compared with the isogenic wild-type Guy11.
Both _alb1__-_ (Supplementary Video 4) and _buf1__-_ mutants displayed homogeneous high fluorescence lifetimes of 3.23 ± 0.063 ns and 3.20 ± 0.056 ns, respectively, similar to values for
non-melanized 4 h appressoria in Guy11 (Fig. 3a,b,e). Furthermore, when Guy11 was exposed to the melanin biosynthesis inhibitor tricyclazole at 3 h compared to an untreated control, we
observed a high fluorescence lifetime of 3.18 ± 0.031 ns, consistent with lifetimes of the melanin-deficient mutants and low appressorium tension (Fig. 3c–e). The mechanoprobe N+-BDP
therefore demonstrates that _alb1__-_ and _buf1__-_ mutants do not generate appressorium turgor, and furthermore, that tension in the appressorial membrane of melanin mutants and
tricyclazole-treated Guy11 is universally low. We also tested whether N+-BDP could detect an increase in membrane tension in non-melanized germ tube tips (Fig. 4a) during polarized growth
compared with the wild type (Fig. 4b). Interestingly, the tips and points of curvature of germ tubes exhibited a lower fluorescence lifetime of 3.84 ± 0.078 ns compared with the subapical
walls of the germ tube that displayed an average higher lifetime of 4.20 ± 0.0773 ns (Fig. 4b). Considering that melanin deposition has not begun at 3 hpi, with enzymes involved in melanin
biosynthesis peaking in expression between 6 hpi and 8 hpi (_ALB1_ MGG_07219, _RSY1_ MGG_05059 and _BUF1_ MGG_02252) (ref. 36) (Fig. 4c), this experiment effectively decouples melanization
from membrane tension, showing that turgor generation in actively growing non-melanized hyphal tips can be revealed by the mechanosensor. INVESTIGATING THE GENETIC CONTROL OF APPRESSORIUM
TURGOR GENERATION WITH N+-BDP To test whether N+-BDP could provide new insight into turgor control in _M. oryzae_, we tested mutants impaired in appressorium function in which the effect on
turgor generation is not known. Septins are required for pathogenicity of _M. oryzae_, regulating F-actin organization in the appressorium, and acting as lateral diffusion barriers for
proteins involved in penetration peg emergence and elongation37. A total of six septins are present in _M. oryzae_, four of which share similarity to core septins identified in yeast, Sep3,
Sep4, Sep5 and Sep6. Recently, very long chain fatty acid biosynthesis has been shown to regulate phosphatidylinositol-phosphate-mediated septin ring formation by recruiting septins to
curved plasma membranes, initiating septin ring formation and subsequent penetration peg emergence38. Septin 5 is a core septin of _M. oryzae_ that forms part of the heteropolymeric septin
ring in the appressorium pore and is essential for plant infection37. We stained a _∆sep5_ mutant with N+-BDP to see whether turgor generation is affected by absence of the septin. This
revealed no significant change in membrane tension and appressorium turgor (2.90 ± 0.059 ns) compared with Guy11 (2.79 ± 0.046 ns) (Fig. 5a,b,d), providing evidence that absence of Sep5 has
no effect on appressorium turgor generation, but instead impairs re-polarization, separating the two processes and demonstrating more clearly the precise function of septins in _M.
oryzae_37. We were also curious to test the _∆nox2_ mutant, because previous work has shown that in the absence of _NOX2_, septins and F-actin do not assemble at the appressorium pore39. In
addition to playing an important role in septin-mediated cytoskeletal re-organization, Nox enzymes are implicated in the chemiosmotic generation of turgor pressure, particularly in mammalian
cells40. Staining the _∆nox2_ mutant with the N+-BDP rotor probe revealed a significant reduction in membrane tension (3.12 ± 0.041 ns) compared with the wild-type Guy11 (2.79 ± 0.046 ns)
(Fig. 5a,c,d), suggesting that absence of the Nox2 NADPH oxidase catalytic subunit does affect turgor generation in the appressorium. We conclude that the mechanoprobe N+-BDP is effective as
a means of screening mutants impaired in appressorium function for a role in turgor pressure generation. THE SLN1 KINASE CONTROLS TURGOR THROUGHOUT APPRESSORIUM DEVELOPMENT The Sln1
histidine-aspartate kinase in _M. oryzae_ acts as a sensor to detect when a critical threshold of turgor has been reached in the appressorium to enable host penetration26. As a consequence
Δ_sln1_ mutants are unable to sense turgor and their appressoria are predicted to have excess turgor pressure and are hyper-melanized26,27. We therefore tested whether the N+-BDP
mechanoprobe could detect aberrant turgor generation in a Δ_sln1_ mutant. First, we used septin–green fluorescent protein (GFP) localization to determine the time at which maximum turgor is
achieved, when a septin ring is formed in the appressorium pore to facilitate re-polarization (Fig. 6a and Supplementary Video 5)8,37,41. F-actin and septin ring recruitment occurs in a
pressure-dependent-manner26,42 while in a melanin-deficient _buf1__-_ mutant12, septin and F-actin localization are disordered26,41,42. The hyper-melanized Δ_sln1_ mutant also displays
aberrant septin and actin localization patterns (Fig. 6c and Supplementary Video 6)26. To investigate whether the N+-BDP could detect the predicted abnormal turgor of Δ_sln1_ mutants we
carried out staining of a time course of infection-related development and determined the average fluorescence lifetime at each developmental stage. In Guy11 appressoria at 4 hpi, an average
lifetime of 3.95 ± 0.091 ns was observed, which significantly reduced to 3.11 ± 0.061 ns at 6 hpi, consistent with initiation of melanin synthesis and onset of turgor generation. By 8 hpi
the average lifetime had significantly reduced again to 2.81 ± 0.079 ns, and by 24 hpi the average lifetime was 2.73 ± 0.042 ns. The average lifetime of N+-BDP fluorescence did not
significantly change between 8 hpi and 24 hpi, suggesting either that membrane tension remains constant after 8 hpi or that the rotor probe is saturated and unable to resolve higher tensions
(Fig. 6b,e). The commitment point for septin ring organization is between 8 h and 10 h (refs. 8,37). Previously, septin ring formation was shown to be impaired after lowering appressorium
turgor by application of exogenous glycerol, or treatment with tricyclazole when applied up to 16 hpi (ref. 26). This suggests that appressorium turgor reaches a critical threshold before
septin ring assembly, and its modulation and maintenance through action of the Sln1-turgor-sensing-complex helps to stabilize conditions required for preserving septin ring organization. In
the Δ_sln1_ mutant we observed significantly higher membrane tension particularly between 4 hpi and 6 hpi, averaging 3.70 ± 0.057 ns and 2.89 ± 0.110 ns, respectively. By 8 hpi the average
lifetime significantly reduced to 2.55 ± 0.057 ns, and by 24 hpi the average lifetime observed was 2.51 ± 0.062 ns. Once again, the lifetime of the N+-BDP rotor probe remained constant
between 8 hpi and 24 hpi, but the lifetime was significantly lower at 4 hpi, 8 hpi and 24 hpi compared with Guy11 (Fig. 6d,e and Supplementary Video 7). N+-BDP therefore reveals that Sln1 is
necessary for controlling the temporal dynamics of turgor generation throughout appressorium development, because the Δ_sln1_ mutant exhibits excess turgor even at 4 hpi. In the current
model for the action of Sln1, it is proposed to act as a negative regulator of turgor once a threshold of turgor has been achieved. The model is therefore likely to be incorrect, or at least
incomplete because it appears likely that the Sln1 kinase orchestrates turgor generation throughout appressorium development, controlling both the rate of turgor build-up and when
sufficient pressure has been generated to facilitate the switch to polarized growth. Live cell imaging with the N+-BDP rotor probe therefore reveals new insight into the regulation of
appressorium turgor generation. DISCUSSION Many fungal pathogens develop infection cells to breach the tough external barrier of a plant or animal1,2. These cells include appressoria,
hyphopodia and infection cushions2,3,28,43,44,45. Appressoria are, however, the most studied infection structure and essential for many of the most destructive plant diseases46. Economically
important pathogens such as the powdery mildew pathogen _Blumeria graminis_, the corn smut fungus _Ustilago maydis_ and soybean rust fungus _Phakopsora pachyrhizi_, for example, all
elaborate appressoria. Oomycete pathogens, such as _Phytophthora_ and _Pythium_ species also develop functional appressoria2, and recently it was shown that the late blight pathogen
_Phytophthora infestans_ enters its host at a diagonal angle, using a specific ‘naifu’ cutting action to break the host leaf surface47. The devastating rice blast fungus _M. oryzae_ uses
appressoria to breach the tough rice leaf cuticle, by generating enormous turgor of up to 8.0 MPa (ref. 13). In this study we applied a plasma membrane targeting rigidochromic molecular
rotor to generate complete mechanical maps of wild-type and mutant appressoria of _M. oryzae_. These have revealed how the mechanics of the plasma membrane are adaptively modulated to
accommodate appressorium growth and turgor generation. Creating detailed tension maps of appressoria during different stages of infection-related development has allowed us to observe
real-time changes in turgor generation. Previous studies have suggested that changes in mechanical tension of a composite lipid membrane are facilitated through formation of bulges and
protrusions of membrane domains48. In addition, other studies have suggested how mechanical stress on a membrane can increase the line tension between a microdomain and the rest of the lipid
bilayer49, which can in turn lead to microdomain growth50. How membranes deal with such extreme tensions, however, is unknown, because all membrane studies have been performed on cells with
much lower internal pressures. By contrast, our study used fungal appressoria that generate pressures that are an order of magnitude higher than any previously analysed. This revealed that
the appressorium membrane develops different microdomains when subject to extreme levels of mechanical stress. Control experiments with a probe sensitive to chemical polarity of membranes
that can reveal compositional inhomogeneities, but that is tension insensitive, showed that the unexpected tension inhomogeneities do not correlate with formation of lipid microdomains or
melanin binding of the probe. This is further corroborated by the fact that N+-BDP can detect turgor changes in the tips of non-melanized hyphae. Our study provides new information regarding
how membrane tension is distributed under extreme turgor in eukaryotic cells, revealing much larger membrane spatial heterogeneities than previously reported. While previous studies in
unwalled cells and free-standing in vitro membranes have linked tension to changes in membrane homogeneity48,50, these are not pressurized to the same extent as membranes enclosed in
appressoria. The findings presented here are therefore the first report of membrane heterogeneity under enormous pressures, and the results highlight how increases in tension can give rise
to regimes in membrane physics, which are unknown and probably reflect the extreme mechanics of an appressorium. One potential explanation for the large tension inhomogeneities is that
tension on the plasma membrane is very sensitive to the state of its anchoring to the cell wall in which it is enclosed, suggesting processes that couple wall and membrane mechanics. There
are numerous proteins, including a variety of receptors, for example, that feature both transmembrane and extracellular domains and that mechanically link the membrane to the cell wall,
potentially in a non-homogeneous manner that may be reflected in the tension patterns revealed. To test the efficacy of the N+-BDP mechanosensor, we first validated the well-known role of
melanin in appressorium turgor generation. Here the rotor dye was able to reveal the severe impairment in appressorium turgor generation and the lack of membrane heterogeneity that
accompanied reduced pressure of _M. oryzae_ melanin-deficient mutants. We also showed that mutants in which turgor dynamics have not been investigated can be readily studied using N+-BDP.
While septin assembly is necessary for appressorium re-polarization and penetration peg emergence, our analysis revealed that a Δ_sep5_ mutant did not show a significant reduction in turgor,
based on FLIM analysis. This is consistent with previous reports that have shown that septin assembly is turgor dependent and occurs only once a threshold of pressure has been generated in
the appressorium26,41. By contrast, the Δ_nox2_ mutant showed a reduction in appressorium turgor. The Nox2 NAPDH oxidase catalytic subunit is necessary for appressorium function, including
septin assembly and penetration peg formation39. Our analysis suggests that Nox2 may act upstream of septin assembly, serving a wider role in appressorium maturation than hitherto
appreciated, including ensuring that sufficient turgor is generated. To investigate whether this is a direct result of inhibiting reactive oxygen species generation, chemical inhibition with
ascorbic acid or the flavocytochrome inhibitor diphenylene iodonium could be carried out. It would also be valuable to investigate the function of the regulatory subunit NoxR in
conditioning the ability of Nox2 to regulate appressorium turgor. Finally, we tested whether N+-BDP could reveal perturbations in appressorium turgor associated with the ∆_sln1_ mutant26.
Sln1 has been proposed to act as a turgor sensor kinase and is necessary to enable a mature appressorium to re-polarize and cause infection26. A mathematical model of appressorium-mediated
plant infection predicted that a mutant lacking such a sensor would be unable to modulate appressorium turgor and therefore display excess pressure, but would be unable to re-polarize26. The
∆_sln1_ mutant displays these phenotypes, but until now its excess turgor was only predicted using the incipient cytorrhysis assay. Here we observed that ∆_sln1_ mutants do show excess
appressorium membrane tension revealed by the N+-BDP rotor. Even though we are clearly operating close to the limit of resolution of the rotor dye based on our calibration curve, because of
the enormous pressures being measured, which are well beyond the scope of pressure probes for instance, a significant difference in turgor can be determined in mature appressoria of ∆_sln1_
mutants compared with Guy11. This provides direct evidence that Sln1 does act as a sensor of appressorium turgor as predicted26, but also shows that the kinase must act earlier than
anticipated because it clearly modulates turgor during the initial stages of appressorium formation. Sln1 is therefore likely to control the rate of turgor generation, in addition to the
switch to polarized growth and septin-dependent plant infection, playing a key role in the orchestration of plant infection. When considered together, these experiments highlight the utility
of the N+-BDP mechanosensor as a direct means of analysing membrane tension in a living appressorium. Fungal mechanobiology is a new and exciting approach for studying the mechanics of the
plasma membrane, with scope to explore other compartments including the fungal cell wall, vacuoles and cytosol24. Future experiments employing the use of the molecular rotor may provide new
quantifiable insights to spatial variations in microviscosity at the point of penetration, and at crossing points during cell-to-cell movement, where it is possible that transpressoria–
which form specifically at cell junctions –generate turgor to successfully breach neighbouring cells3,51. Furthermore, a combination of surface-deformation imaging, rotor probe staining and
mathematical modelling could help to establish the precise mode of entry and the forces translated at the host leaf surface which will prove invaluable in the search for effective blast
disease control strategies47. METHODS FUNGAL STRAINS AND GROWTH CONDITIONS Growth and maintenance of _M. oryzae_ was performed as described previously52. All strains used in the study are
stored in the laboratory of N.J.T. and are freely available upon request. TWO-DIMENSIONAL FLIM IMAGING Appressorium development was induced in vitro on borosilicate 18 × 22 mm glass
coverslips (Thermo Fisher Scientific), as described previously53. For onion epidermis experiments, epidermal layers were incubated in chloroform for 30 min, washed five times in sterile
dH2O, before mounting onto a coverslip for inoculation. A 50 μl aliquot of conidial suspension (5 × 104 ml−1) was placed on a coverslip/onion epidermis and incubated at 24 °C. The aqueous
phase of the droplet from Guy11 2–3 h germ tubes, 4 h, 24 h and _C. higginsianum_ early and late appressoria was replaced with a 50 μl droplet of 10 μmol l−1 N+-BDP probe dissolved in
sterile distilled H2O. Staining was performed at room temperature for 20 min for the hyper-melanized Δ_sln1_ mutant and for 5 min for the wild-type Guy11 and all other mutants, after which
unbound dye was removed by replacing 50 μl of the droplet five times with water. For calibration of the probe in appressoria, the aqueous phase of the droplet was removed and replaced with a
50 μl droplet of glycerol (0.2 M, 0.4 M, 0.75 M or 1 M). FLIM imaging was performed on a Leica TCS SP8X upright scanning confocal microscope coupled to a PicoHarp 300 TCSPC module
(PicoQuant GmbH). Samples were excited with 488 nm output of a pulsed SuperK EXTREME supercontinuum white light laser (NKT Photonics) working at a repetition rate of 20 MHz. The full width
at half maximum of the laser pulse was ~170 ps, as determined from instrument response functions recorded using Erythrosin B (Sigma-Aldrich, >95% purity) in KI-saturated (Sigma-Aldrich)
water. Fluorescence emission was captured in the 510–530 nm range using a Leica HyD SMD detector. The objective lens was an HC Plan Apo 63×/numerical aperture 1.20 water immersion objective
(Leica Microsystems). SymPhoTime 64 (version 2.4, PicoQuant GmbH) was used to draw by hand in the 2D image a region of interest (ROI) that closely followed the membrane of the appressorium
and did not include signal arising from any other part of the sample54. The same software was used to fit the overall fluorescence decay curve of the ROI with a three-component exponential
decay function. Fits were deemed acceptable only if residuals were evenly distributed around zero and the _χ_2 values were within 0.70–1.30 range. The average fluorescence lifetimes reported
in this work are the intensity-weighted average lifetimes, which have been calculated as $$< \tau > =\frac{\sum _{i}{\alpha }_{i}{\tau }_{i}^{2}}{\sum _{i}{\alpha }_{i}{\tau }_{i}}$$
where _a__i_ and _t__i_ are the amplitude and the lifetime of species _i_, respectively55. Images are reported in a false-colour scale that represents the weighted average of fluorescence
decay for each pixel. For multi-exponential decays, the weighted average of fluorescence decay is equivalent to the intensity-weighted average fluorescence lifetime. Each datapoint in our
analysis corresponds to the weighted average fluorescence lifetime obtained for the ROI of a single 2D image of an individual appressorium. FLIM TIME SERIES EXPERIMENTS Conidia were
collected and inoculated onto glass coverslips. Early appressoria at 4 hpi were stained, washed and mounted onto glass slides, as previously described. FLIM imaging was performed on a
Stellaris 8 Falcon upright scanning confocal microscope (Leica Microsystems). Samples were excited at 488 nm using a pulsed SuperK Fianium FIB-12 PP laser source (NK Photonics) working at a
20 MHz repetition rate. The full width at half maximum of the laser pulse was ~190 ps, as determined from instrument response functions obtained using Erythrosin B solution. Detection range
and objective lens were the same as described above. A Leica HyD X detector was used and images acquired at 0 (4.5 h appressoria), 13, 31, 48, 65, 83, 109, 125, 138 and 146 min (7.5 h
appressoria). The images were processed and analysed in LAS X (version 4.2, Leica Microsystems) and movies generated using Python distributed under the GNU public licence (Anaconda Ver.
3.8.10., https://anaconda.com). Python libraries used to generate the movie were NumPy56, Scikit-image57, pystackreg58 and OpenCV59. The Python script can be found at
https://github.com/SergioGabrielLopez/movie_script. The fluorescence lifetime for each frame was obtained by selecting an ROI and then fitting the overall fluorescence decay of the ROI with
a four-exponential decay function. The fit was judged according to the previously mentioned criteria. THREE-DIMENSIONAL LIFETIME IMAGING _M. oryzae_ conidia were inoculated onto glass
coverslips, stained with N+-BDP and imaged at desired times. Images were acquired on a Stellaris 8 FALCON upright scanning confocal microscope (Leica Microsystems). All imaging parameters
were identical to those described for acquisition of the FLIM time series. The _z_-stacks had a length in the _z_-direction of ~12–15 mm, took 3–7 min to complete and were acquired in
compliance with the Nyquist-Shannon sampling theorem. The 3D rendering of the _z_-stacks was carried out in LAS X (version 4.2, Leica Microsystems). 2D IMAGING OF APPRESSORIA USING NR12S
CHEMICAL POLARITY PROBE To image plasma membrane polarity in _M. oryzae_ appressoria using the chemical polarity probe NR12S, a portion of the aqueous phase of the droplet (50 μl), was
replaced with a solution of NR12S, dissolved at 10 μmol l−1 in water. Staining was performed for 7 min, after which any unbound dye was removed by replacing 50 μl of the droplet five times
with water. Two-dimensional ratiometric imaging with NR12S was performed on a Leica TCS SP8X upright scanning confocal microscope. Samples were excited with 514 nm output of a SuperK EXTREME
supercontinuum white light laser (NKT Photonics) working at repetition rate of 80 MHz. Fluorescence was detected at 529–585 (‘blue channel’) and 610–700 nm (‘red channel’) using Leica HyD
SMD detectors. Ratiometric images obtained with NR12S staining were constructed from recorded intensity images using a custom MATLAB routine that divides photon count in each pixel of the
blue channel image, by photon count in the corresponding pixel in the red channel image. Resulting images are reported in a false-colour scale that represents the intensity ratio for each
pixel. REPORTING SUMMARY Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY The _M. oryzae_ genome
database used in this study was http://fungi.ensembl.org/Magnaporthe_oryzae/Info/Index. All _M. oryzae_ strains used in this study are freely available upon request from the corresponding
authors. Detailed protocols for synthesis of both the N+-BDP rotor probe and the chemical polarity probe NR125 are available on request from J.S. Source data are provided with this paper.
CODE AVAILABILITY The Python script used to produce the _M. oryzae_ FLIM movie has been publicly deposited in GitHub at https://github.com/SergioGabrielLopez/movie_script. REFERENCES *
Mendgen, K., Hahn, M. & Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. _Annu. Rev. Phytopathol._ 34, 367–386 (1996). CAS PubMed Google Scholar *
Talbot, N. J. Appressoria. _Curr. Biol._ 29, R144–R146 (2019). CAS PubMed Google Scholar * Ryder, L. S. et al. The appressorium at a glance. _J. Cell Sci._
https://doi.org/10.1242/jcs.259857 (2022). * Brown, W. & Harvey, C. Studies in the physiology of parasitism. X. On the entrance of parasitic fungi into the host plant. _Ann. Bot._ 41,
643–662 (1927). Google Scholar * Zhang, N. et al. Generic names in Magnaporthales. _IMA Fungus_ 7, 155–159 (2016). PubMed PubMed Central Google Scholar * Fisher, M. C. et al. Emerging
fungal threats to animal, plant and ecosystem health. _Nature_ 484, 186–194 (2012). PubMed Google Scholar * Wilson, R. A. & Talbot, N. J. Under pressure: investigating the biology of
plant infection by _Magnaporthe oryzae_. _Nat. Rev. Microbiol._ 7, 185 (2009). CAS PubMed Google Scholar * Eseola, A. B. et al. Investigating the cell and developmental biology of plant
infection by the rice blast fungus _Magnaporthe oryzae_. _Fungal Genet. Biol._ 154, 103562 (2021). CAS PubMed Google Scholar * de Jong, J. C., McCormack, B. J., Smirnoff, N. & Talbot,
N. J. Glycerol generates turgor in rice blast. _Nature_ 389, 244–244 (1997). Google Scholar * Bechinger, C. et al. Optical measurements of invasive forces exerted by appressoria of a plant
pathogenic fungus. _Science_ 285, 1896–1899 (1999). CAS PubMed Google Scholar * Money, N. P. & Harold, F. M. Two water molds can grow without measurable turgor pressure. _Planta_
190, 426–430 (1993). Google Scholar * Chumley, F. G. & Valent, B. Genetic analysis of melanin-deficient, nonpathogenic mutants of _Magnaporthe grisea_. _Mol. Plant Microbe_ 3, 135–143
(1990). CAS Google Scholar * Talbot, N. J. On the trail of a cereal killer: exploring the biology of _Magnaporthe grisea_. _Annu. Rev. Microbiol._ 57, 177–202 (2003). CAS PubMed Google
Scholar * Foster, A. J., Ryder, L. S., Kershaw, M. J. & Talbot, N. J. The role of glycerol in the pathogenic lifestyle of the rice blast fungus _Magnaporthe oryzae_. _Environ.
Microbiol._ 19, 1008–1016 (2017). PubMed Google Scholar * Howard, R. J., Ferrari, M. A., Roach, D. H. & Money, N. P. Penetration of hard substrates by a fungus employing enormous
turgor pressures. _Proc. Natl Acad. Sci. USA_ 88, 11281–11284 (1991). CAS PubMed PubMed Central Google Scholar * Riggi, M. et al. Decrease in plasma membrane tension triggers
PtdIns(4,5)P(2) phase separation to inactivate TORC2. _Nat. Cell Biol._ 20, 1043–1051 (2018). CAS PubMed PubMed Central Google Scholar * Sun, G., Elowsky, C., Li, G. & Wilson, R. A.
TOR-autophagy branch signaling via Imp1 dictates plant-microbe biotrophic interface longevity. _PLoS Genet._ 14, e1007814–e1007814 (2018). PubMed PubMed Central Google Scholar *
Lopez-Berges, M. S., Rispail, N., Prados-Rosales, R. C. & Di Pietro, A. A nitrogen response pathway regulates virulence functions in _Fusarium oxysporum_ via the protein kinase TOR and
the bZIP protein MeaB. _Plant Cell_ 22, 2459–2475 (2010). CAS PubMed PubMed Central Google Scholar * Marroquin-Guzman, M., Sun, G. C. & Wilson, R. A. Glucose-ABL1-TOR signaling
modulates cell cycle tuning to control terminal appressorial cell differentiation. _PLoS Genet_. https://doi.org/10.1371/journal.pgen.1006557 (2017). * Oh, Y. et al. Transcriptome analysis
reveals new insight into appressorium formation and function in the rice blast fungus Magnaporthe oryzae. _Genome Biol._ 9, R85 (2008). PubMed PubMed Central Google Scholar * Qian, B. et
al. MoPpe1 partners with MoSap1 to mediate TOR and cell wall integrity signalling in growth and pathogenicity of the rice blast fungus _Magnaporthe oryzae_. _Environ. Microbiol._ 20,
3964–3979 (2018). CAS PubMed PubMed Central Google Scholar * Yu, F. W. et al. The TOR signaling pathway regulates vegetative development and virulence in _Fusarium graminearum_. _N.
Phytol._ 203, 219–232 (2014). CAS Google Scholar * Zhu, X. M. et al. A VASt-domain protein regulates autophagy, membrane tension, and sterol homeostasis in rice blast fungus. _Autophagy_
17, 2939–2961 (2021). CAS PubMed Google Scholar * Michels, L. et al. Complete microviscosity maps of living plant cells and tissues with a toolbox of targeting mechanoprobes. _Proc. Natl
Acad. Sci. USA_ 117, 18110–18118 (2020). CAS PubMed PubMed Central Google Scholar * Colom, A. et al. A fluorescent membrane tension probe. _Nat. Chem._ 10, 1118–1125 (2018). CAS PubMed
PubMed Central Google Scholar * Ryder, L. S. et al. A sensor kinase controls turgor-driven plant infection by the rice blast fungus. _Nature_ 574, 423–427 (2019). CAS PubMed Google
Scholar * Zhang, H. et al. A two-component histidine kinase, MoSLN1, is required for cell wall integrity and pathogenicity of the rice blast fungus, Magnaporthe oryzae. _Curr Genet_ 56,
517–528 (2010). CAS PubMed Google Scholar * Ryder, L. S. & Talbot, N. J. Regulation of appressorium development in pathogenic fungi. _Curr. Opin. Plant Biol._ 26, 8–13 (2015). CAS
PubMed PubMed Central Google Scholar * Rocha, R. O., Elowsky, C., Pham, N. T. T. & Wilson, R. A. Spermine-mediated tight sealing of the _Magnaporthe oryzae_ appressorial pore–rice
leaf surface interface. _Nat. Microbiol_ 5, 1472–1480 (2020). CAS PubMed Google Scholar * Lingwood, D. & Simons, K. Lipid rafts as a membrane-organizing principle. _Science_ 327,
46–50 (2010). CAS PubMed Google Scholar * Silvius, J. R. Role of cholesterol in lipid raft formation: lessons from lipid model systems. _Biochim. Biophys. Acta_ 1610, 174–183 (2003). CAS
PubMed Google Scholar * Brown, D. A. & London, E. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. _J. Biol. Chem._ 275, 17221–17224 (2000). CAS PubMed
Google Scholar * Yan, Y. et al. _Colletotrichum higginsianum_ as a model for understanding host–pathogen interactions: a review. _Int. J. Mol. Sci._ 19, 2142 (2018). PubMed PubMed Central
Google Scholar * Kucherak, O. A. et al. Switchable Nile red-based probe for cholesterol and lipid order at the outer leaflet of biomembranes. _J. Am. Chem. Soc._ 132, 4907–4916 (2010).
CAS PubMed Google Scholar * Michels, L. et al. Molecular sensors reveal the mechano-chemical response of _Phytophthora infestans_ walls and membranes to mechanical and chemical stress.
_Cell Surf._ 8, 100071 (2022). CAS PubMed PubMed Central Google Scholar * Oses-Ruiz, M. et al. Appressorium-mediated plant infection by _Magnaporthe oryzae_ is regulated by a
Pmk1-dependent hierarchical transcriptional network. _Nat. Microbiol._ 6, 1383–U1106 (2021). CAS PubMed Google Scholar * Dagdas, Y. F. et al. Septin-mediated plant cell invasion by the
rice blast fungus, _Magnaporthe oryzae_. _Science_ 336, 1590–1595 (2012). CAS PubMed Google Scholar * He, M. et al. Discovery of broad-spectrum fungicides that block septin-dependent
infection processes of pathogenic fungi. _Nat. Microbiol_ 5, 1565–1575 (2020). CAS PubMed Google Scholar * Ryder, L. S. et al. NADPH oxidases regulate septin-mediated cytoskeletal
remodeling during plant infection by the rice blast fungus. _Proc. Natl Acad. Sci. USA_ 110, 3179 (2013). CAS PubMed PubMed Central Google Scholar * Segal, A. W. NADPH oxidases as
electrochemical generators to produce ion fluxes and turgor in fungi, plants and humans. _Open Biol._ 6, 160028 (2016). PubMed PubMed Central Google Scholar * Dulal, N., Rogers, A., Wang,
Y. & Egan, M. Dynamic assembly of a higher-order septin structure during appressorium morphogenesis by the rice blast fungus. _Fungal Genet. Biol._ 140, 103385 (2020). CAS PubMed
Google Scholar * Dulal, N. et al. Turgor-dependent and coronin-mediated F-actin dynamics drive septin disc-to-ring remodeling in the blast fungus _Magnaporthe oryzae_. _J. Cell Sci._ 134,
jcs251298 (2021). CAS PubMed Google Scholar * Goos, R. D. & Gessner, R. V. Hyphal modifications of _Sphaerulina pedicellata_—Appressoria or Hyphopodia. _Mycologia_ 67, 1035–1038
(1975). Google Scholar * Bozkurt, T. O. & Kamoun, S. The plant–pathogen haustorial interface at a glance. _J Cell Sci_ https://doi.org/10.1242/jcs.237958 (2020). * Choquer, M. et al.
The infection cushion of _Botrytis cinerea_: a fungal ‘weapon’ of plant-biomass destruction. _Environ. Microbiol._ 23, 2293–2314 (2021). CAS PubMed Google Scholar * Dean, R. et al. The
top 10 fungal pathogens in molecular plant pathology. _Mol. Plant Pathol._ 13, 414–430 (2012). PubMed PubMed Central Google Scholar * Bronkhorst, J. et al. A slicing mechanism facilitates
host entry by plant-pathogenic _Phytophthora_. _Nat. Microbiol._ 6, 1000–1006 (2021). CAS PubMed Google Scholar * Sens, P. & Turner, M. S. Budded membrane microdomains as tension
regulators. _Phys. Rev. E_ 73, 031918 (2006). Google Scholar * Akimov, S. A., Kuzmin, P. I., Zimmerberg, J. & Cohen, F. S. Lateral tension increases the line tension between two domains
in a lipid bilayer membrane. _Phys. Rev. E_ 75, 011919 (2007). Google Scholar * García-Sáez, A. J., Chiantia, S. & Schwille, P. Effect of line tension on the lateral organization of
lipid membranes. _J. Biol. Chem._ 282, 33537–33544 (2007). PubMed Google Scholar * Cruz-Mireles, N., Eseola, A. B., Oses-Ruiz, M., Ryder, L. S. & Talbot, N. J. From appressorium to
transpressorium—defining the morphogenetic basis of host cell invasion by the rice blast fungus. _PLoS Pathog_. https://doi.org/10.1371/journal.ppat.1009779 (2021). * Talbot, N. J., Ebbole,
D. J. & Hamer, J. E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus _Magnaporthe grisea_. _Plant Cell_ 5, 1575–1590 (1993). CAS
PubMed PubMed Central Google Scholar * Hamer, J. E., Howard, R. J., Chumley, F. G. & Valent, B. A mechanism for surface attachment in spores of a plant pathogenic fungus. _Science_
239, 288 (1988). CAS PubMed Google Scholar * Kagan, V. E. et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. _Nat. Chem. Biol._ 13, 81–90 (2017). CAS PubMed
Google Scholar * Li, Y. et al. Investigations on average fluorescence lifetimes for visualizing multi-exponential decays. Front. Phys. https://doi.org/10.3389/fphy.2020.576862 (2020). *
Harris, C. R. et al. Array programming with NumPy. _Nature_ 585, 357–362 (2020). CAS PubMed PubMed Central Google Scholar * van der Walt, S. et al. scikit-image: image processing in
Python. _PeerJ_ 2, e453 (2014). PubMed PubMed Central Google Scholar * Thévenaz, P., Ruttimann, U. E. & Unser, M. A pyramid approach to subpixel registration based on intensity. _IEEE
Trans. Image Process._ 7, 27–41 (1998). PubMed Google Scholar * Pulli, K., Baksheev, A., Kornyakov, K. & Eruhimov, V. Realtime computer vision with OpenCV: mobile computer-vision
technology will soon become as ubiquitous as touch interfaces. _Queue_ 10, 40–56 (2012). Google Scholar Download references ACKNOWLEDGEMENTS This project was funded by the BBSRC grant
BB/V016342/1 and by the Gatsby Charitable Foundation. We thank C. Faulkner for providing us with _C. higginsianum_ strain IMI 349061. We thank C. Wood for critical reading of the manuscript.
AUTHOR INFORMATION Author notes * These authors contributed equally: Sergio G. Lopez, Lucile Michels. AUTHORS AND AFFILIATIONS * The Sainsbury Laboratory, University of East Anglia, Norwich
Research Park, Norwich, UK Lauren S. Ryder, Alice B. Eseola, Weibin Ma & Nicholas J. Talbot * Cell and Developmental Biology, The John Innes Centre, Norwich Research Park, Norwich, UK
Sergio G. Lopez * Laboratory of Biochemistry, Wageningen University & Research, Wageningen, the Netherlands Lucile Michels & Joris Sprakel Authors * Lauren S. Ryder View author
publications You can also search for this author inPubMed Google Scholar * Sergio G. Lopez View author publications You can also search for this author inPubMed Google Scholar * Lucile
Michels View author publications You can also search for this author inPubMed Google Scholar * Alice B. Eseola View author publications You can also search for this author inPubMed Google
Scholar * Joris Sprakel View author publications You can also search for this author inPubMed Google Scholar * Weibin Ma View author publications You can also search for this author inPubMed
Google Scholar * Nicholas J. Talbot View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS L.S.R, N.J.T. and J.S. conceptualized the project.
Experimental analyses were carried out by L.S.R, S.G.L, L.M, A.B.E and W.M. The paper was written by L.S.R, J.S and N.J.T. CORRESPONDING AUTHOR Correspondence to Nicholas J. Talbot. ETHICS
DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Nature Microbiology_ thanks Ashley Garrill, Bart Thomma and the other,
anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral
with regard to jurisdictional claims in published maps and institutional affiliations. EXTENDED DATA EXTENDED DATA FIG. 1 REGION OF INTEREST SELECTION IN TWO DIMENSIONAL FLIM IMAGES. A,
Representative FLIM image of a wild type Guy11 appressorium stained with the N+-BDP probe at 24 h. B, Image a showing the region of interest in colour with all other pixels in grayscale.
EXTENDED DATA FIG. 2 SPATIAL VARIATIONS IN MEMBRANE TENSION IN GERM TUBES AND APPRESSORIA OF _M. ORYZAE_. Representative FLIM images of wild type Guy11 24 h rotor stained appressoria and
germ tubes. Images are representative of _n_ = _3_ independent repeats of the experiment. Scale bar= 10 μm. EXTENDED DATA FIG. 3 ROTOR DYE N+-BDP CALIBRATION IN _M. ORYZAE_ APPRESSORIA
INCUBATED IN GLYCEROL. Line graph showing the average fluorescence lifetime of N+-BDP stained appressoria of Guy11 incubated in different molar concentrations of glycerol. Values are means ±
SEM for 3 biological replicates of the experiment, total observations _n_ = 129 appressoria in 3 biological replicates; typical range 22–34 appressoria; data are presented as mean ± SEM.
***_P_ < 0.0002 as determined by one-way analysis (ANOVA) with Dunnett’s multiple comparisons test. Source data EXTENDED DATA FIG. 4 MEMBRANE TENSION OF THE APPRESSORIUM DECREASES DURING
MATURATION. Guy11 appressoria were stained with the rotor probe N+-BDP at 4.5 hpi (0 min) and FLIM images captured for 3 h and fluorescence lifetimes plotted. Source data EXTENDED DATA FIG.
5 THE MECHANOSENSOR N+-BDP REVEALS SPATIAL VARIATIONS IN MEMBRANE TENSION IN _M. ORYZAE_ APPRESSORIA ON A YIELDING PLANT SURFACE. A, Time course of _M. oryzae_ infection-related-development
on onion epidermis treated with chloroform. Images are representative of _n_ = _3_ independent repeats of the experiment. B, FLIM micrographs of Guy11 at 24 h germinated on onion epidermis
treated with chloroform and stained with the rotor probe N+-BDP. The colour corresponds to the fluorescence lifetime values expressed in nanoseconds, as shown in the key 2–4 ns. C, Dot plot
showing the average fluorescence lifetime for 24hpi Guy11 appressoria developed on onion epidermis. Each dot corresponds to the average fluorescence lifetime obtained for an ROI drawn around
the membrane of an individual appressorium in a 2D FLIM image. Total observations _n_ = 21 appressoria in 3 biological replicates; each biological replicate is colour coded (R1: replicate
1, R2: replicate 2, R3: replicate 3); data are presented as mean ± SEM. Source data EXTENDED DATA FIG. 6 MAPPING SPATIAL VARIATION IN CHEMICAL POLARITY OF THE PLASMA MEMBRANE IN _M. ORYZAE_
APPRESSORIA USING THE SOLVATOCHROMIC PROBE NR12S. A, Chemical structure of NR12S. B,C Schematic illustrations showing the mechanism by which NR12S reports changes in chemical polarity and
lipid phase. D-J, Intensity ratio chemical polarity maps in _M. oryzae_ wild type strain Guy11 4 h appressoria. Images E, F, H and I are magnified areas of 4 h appressoria. The colour scale
translates the intensity ratio values (_n_ = 31 three independent repetitions of the experiment were performed). N+-BDP 4 h and N+-BDP 24 h FLIM images included for comparison. K-P,
Intensity ratio chemical polarity maps of _M. oryzae_ wild type strain Guy11 24 h appressoria. Images I, M, O and P are zoomed in areas of 24 h appressoria, Total observations _n_ = 34
appressoria in 3 biological replicates. Images are representative of _n_ = _3_ independent repeats of the experiment. Images D, G, J, K and N, scale bars = 5 μm, images E, F, H, I, L, M, O
and P scale bars= 1 μm. Image created with BioRender.com. SUPPLEMENTARY INFORMATION REPORTING SUMMARY PEER REVIEW FILE SUPPLEMENTARY VIDEO 1 THREE-DIMENSIONAL FLIM ROTATIONAL MOVIE OF GUY11
4 H APPRESSORIUM. Three-dimensional FLIM of N+-BDP stained 4 h incipient appressoria of Guy11 was performed on a Stellaris 8 FALCON upright scanning confocal microscope. Conidia were
collected from the _M. oryzae_ wild-type Guy11 and inoculated onto glass coverslips. SUPPLEMENTARY VIDEO 2 THREE-DIMENSIONAL FLIM ROTATIONAL MOVIE OF GUY11 7.5 H APPRESSORIUM.
Three-dimensional FLIM of N+-BDP stained Guy11 appressoria at 7.5 h was performed on a Stellaris 8 FALCON upright scanning confocal microscope. Conidia were collected from the _M. oryzae_
wild-type strain Guy11 and inoculated onto glass coverslips. SUPPLEMENTARY VIDEO 3 LIVE CELL IMAGING OF TURGOR GENERATION IN A DEVELOPING GUY11 APPRESSORIUM STAINED WITH THE ROTOR PROBE
N+-BDP 4.5 H 7 HPI. Conidia were collected from a _M. oryzae_ Guy11 and inoculated on glass coverslips. The 4 h appressoria were incubated with 80 µL of N+-BDP rotor probe at 10 µmol−1 in
water for 5 min, washed five times and imaged. SUPPLEMENTARY VIDEO 4 THREE-DIMENSIONAL FLIM ROTATIONAL MOVIE OF A 24 H APPRESSORIUM OF THE _ALB1_ MELANIN-DEFICIENT MUTANT. Three-dimensional
FLIM of _alb1_ 24 h appressoria stained with N+-BDP performed on a Stellaris 8 FALCON upright scanning confocal microscope. Conidia were collected from a _M. oryzae_ wild-type Guy11 and
inoculated onto glass coverslips. SUPPLEMENTARY VIDEO 5 DYNAMIC ASSEMBLY OF A SEPTIN RING IN GUY11 APPRESSORIA. Live cell imaging of septin dynamics during appressorium development in _M.
oryzae_. Movie shows Guy11 expressing Sep5–GFP during infection-related development on a hydrophobic glass coverslip. The movie was captured using a Leica SP8 laser confocal microscope 0–24
h. The movie is a maximum projection _z_-stack. Frames were captured every 5 min and are displayed at 15 frames s−1. Time scale is in hours:minutes:seconds. Scale bar, 5 μm. SUPPLEMENTARY
VIDEO 6 ABERRANT SEPTIN RING AGGREGATION AND HYPER-MELANIZATION IN THE _∆SLN1_ MUTANT. Live cell imaging of aberrant septin dynamics during appressorium development in _M. oryzae_. Movie
shows the _∆sln1_ mutant expressing Sep5–GFP during infection-related development on hydrophobic glass coverslips. The movie was captured using a Leica SP8 laser confocal microscope 0–24
hpi. The movie is a maximum projection _z_-stack. Frames were captured every 5 min and are displayed at 15 frames s−1. Time scale is in hours:minutes:seconds. Scale bar, 10 μm. SUPPLEMENTARY
VIDEO 7 THREE-DIMENSIONAL FLIM ROTATIONAL MOVIE OF _∆SLN1_ 24 H APPRESSORIUM Three-dimensional FLIM of N+-BDP stained _∆sln1_ 24 h appressoria was performed on a Stellaris 8 FALCON upright
scanning confocal microscope. Conidia were collected from _M. oryzae_ wild-type Guy11 and inoculated onto glass coverslips. SOURCE DATA SOURCE DATA FIG. 1 Source Data Fig. 1. SOURCE DATA
FIG. 2 Source Data Fig. 2. SOURCE DATA FIG. 3 Source Data Fig. 3. SOURCE DATA FIG. 4 Source Data Fig. 4. SOURCE DATA FIG. 5 Source Data Fig. 5. SOURCE DATA FIG. 6 Source Data Fig. 6. SOURCE
DATA EXTENDED DATA FIG. 3 Source Data Extended Data Fig. 3. SOURCE DATA EXTENDED DATA FIG. 4 Source Data Extended Data Fig. 4. SOURCE DATA EXTENDED DATA FIG. 5 Source Data Extended Data Fig.
5. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and
reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes
were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If
material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS
ARTICLE Ryder, L.S., Lopez, S.G., Michels, L. _et al._ A molecular mechanosensor for real-time visualization of appressorium membrane tension in _Magnaporthe oryzae_. _Nat Microbiol_ 8,
1508–1519 (2023). https://doi.org/10.1038/s41564-023-01430-x Download citation * Received: 30 August 2022 * Accepted: 19 June 2023 * Published: 20 July 2023 * Issue Date: August 2023 * DOI:
https://doi.org/10.1038/s41564-023-01430-x SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative