Play all audios:
ABSTRACT Complex learned behaviors, like bird song and human speech, develop under the influence of both genetic and environmental factors. Accordingly, learned behaviors comprise species
specificity and individual variability. Auditory information plays a critical role in vocal learning by songbirds, both to memorize tutor songs and to monitor own vocalizations.
Nevertheless, audition-deprived songbirds develop structured, species-specific song patterns. It remains to be elucidated how the auditory input contributes to the development of individual
variability of song characteristics. Here we show that an open-ended vocal learner, the canary, annually recapitulates individually unique songs without audition. Although the total number
of syllable types was reduced by auditory deprivation, other vocal phenotypes examined in the syllable, phrase, and syntax of songs were conserved between the 1st and 2nd years, both in
deafened and intact birds. In deafened canaries, approximately 60% of the syllables were yearly reproduced with consistent acoustic features, whereas the remaining syllables were replaced
with new ones in an annual cycle of song development. These results indicate that the open-ended vocal learning of canaries involves an audition-independent mechanism for the development of
recurrent song idiosyncrasy. SIMILAR CONTENT BEING VIEWED BY OTHERS BALANCED IMITATION SUSTAINS SONG CULTURE IN ZEBRA FINCHES Article Open access 07 May 2021 A MEMORY-DRIVEN AUDITORY PROGRAM
ENSURES SELECTIVE AND PRECISE VOCAL IMITATION IN ZEBRA FINCHES Article Open access 13 September 2021 SONG PREFERENCES PREDICT THE QUALITY OF VOCAL LEARNING IN ZEBRA FINCHES Article Open
access 12 January 2023 INTRODUCTION Birdsong, like human speech, is a form of learned vocalization1,2, which serves two important communicative functions: to signal species specificity and
individual identity3,4,5,6. Songbirds learn syllable acoustics and sequential features from conspecific tutor songs7. Therefore, audition is a crucial sensory input to develop functional
vocalizations, memorize the tutor song, and monitor own vocal outputs. However, juvenile songbirds do not develop their songs from a “blank slate”-like state of extensive plasticity8. Even
when deprived of audition before hearing tutor songs, juvenile birds later develop species-specific songs with a certain degree of structure and individual uniqueness9,10,11,12,13,14.
Previous studies suggest that an intrinsic program _per se_ ensures an effective competency for self-differentiation of structured song patterns. However, it remains to be understood how the
individual uniqueness of specific song features is generated independently from audition and alongside the maintenance of species-specificity. We investigated this question in an open-ended
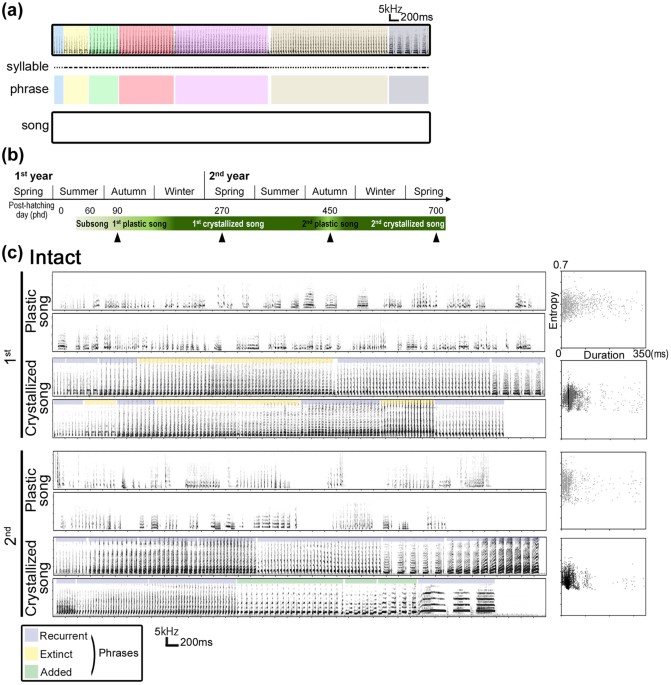
vocal learner, the canary, which annually acquires a set of new song elements, even throughout adulthood15,16,17,18. The song development by the canary have a hierarchical species-specific
structure, composed of syllables and phrases (_i.e_., clusters of repetitive syllables)19 (Fig. 1a). During the 1st post-hatching year, juvenile males start singing subsong in the summer.
The song pattern is crystallized until late winter, and the birds continue to sing it until the following summer (Fig. 1b,c). By the fall of the 2nd year, their songs deteriorate to a degree
of subsong/early-plastic song form, characterized by acoustically amorphous syllables and unstable sequences, after which new songs are re-crystallized by the 2nd year’s late winter20,21.
During the annual song development, three categorical types of syllables, which we termed recurrent, extinct, and added syllables, are identifiable by comparing the crystallized song of the
1st and 2nd years. Taking advantage of this annual syllable subset exchange, we examined whether and how song syllables were recurrent/replaced without auditory input, to investigate the
developmental recurrence of species specificity and individual uniqueness of complex vocalizations in the same individual. In the present study, we deafened individuals of an inbred canary
strain, the Belgian Waterslager, at an early developmental stage during the 1st post-hatching year, and recorded their songs until song crystallization in the 2nd year. We examined the
similarities and differences in the hierarchical structures of their songs between the 1st and 2nd years and between intact and deafened birds. Despite drastic changes in the syllable
phrasing during the annual cycle of song development, deafened canaries iteratively generated species-specific songs with consistent individual uniqueness. This result reveals the existence
of an auditory-independent innate program to generate and retain individual uniqueness within a species-specific song pattern. RESULTS ANNUALLY REPEATED SONG CRYSTALLIZATION AND
DETERIORATION IN THE DEAFENED CANARY To examine the effects of auditory deprivation on the annual development and retention of the individually unique canary songs, juvenile males were
deafened through bilateral cochleae removal, before fledging at 27–34 post-hatching day (phd). This procedure ensured the loss of auditory experience, depriving the canaries from both the
tutor song and their own vocal output. As previously reported13,14, auditory deprivation significantly affected the acquisition of syllable acoustic features in the used canary strain (Fig.
2a–c). However, the annual song development, that is, the timing of song crystallization and successive deterioration, was similarly regulated in both deafened and intact birds. Both
deafened and intact canaries produced plastic songs by the fall and crystallized them from winter to spring (Figs 1c and 2a). Moreover, the crystallized song patterns of both deafened and
intact birds consisted of a series of repertoires with phrases composed of repetitive syllables. However, the crystallized songs of deafened birds included approximately half the number of
syllable types than those of intact birds both for the 1st and 2nd years (_P__*_ = 0.011 and 0.004 in the 1st and 2nd years, respectively; Student’s _t_-test with Holm’s correction) (Fig.
2d), although no significant differences were found between the 1st and 2nd years and between deafened and intact canaries regarding song bout duration and the number of syllables or phrases
in a song (Fig. 2e–g). These results indicate that the annual timings of song crystallization and deterioration were regulated independently from the auditory input. Furthermore, the
fundamental structure of the canary song was also constructed in an auditory-independent way. RECURRENT INTRINSIC UNIQUE SYLLABLE PHRASING IN DEAFENED CANARY We then compared the acoustic
and temporal features of the syllables and phrases in crystallized songs, between the 1st and 2nd years and between intact and deafened birds. On the basis of the acoustic features and
inter-syllable gap timing shown in the spectrogram, we identified three types of syllables, present in the songs of both deafened and intact canaries: extinct, recurrent, and added syllables
(Fig. 3a). Individual differences in the acoustic features of recurrent syllables were maintained and reproduced yearly without auditory input in both deafened and intact birds (Fig. 3b).
After song deterioration in the 2nd year, both deafened and intact birds recapitulated approximately 60% of the individually unique syllables as recurrent syllables in their new crystallized
songs (Fig. 3c,d). This indicates that individually unique syllable variations are retained in an auditory-independent manner. The frequency of occurrence of extinct, recurrent, and added
syllables was conserved between intact and deafened birds (Fig. 3d). In addition, during annual song development, the syllable repertoires were reassembled through the loss of some syllables
(extinct syllables) and addition of new ones (added syllables), resulting in the maintenance of a similar proportion (approximately 30–40%) in songs between the 1st and 2nd years, even in
deafened birds (Fig. 3d). These results indicate that an intrinsic, audition-independent genetic program is necessary and sufficient to regulate syllable maintenance and replacement.
Principal component analysis (PCA) using five acoustic syllable parameters: duration, inter-syllable interval, fundamental frequency, bandwidth, and entropy, revealed the absence of a clear
boundary between the three syllable categories (Fig. 3e). The acoustic features of the syllables in the PCA were mostly merged between intact and deafened birds. In line with this finding,
the syllable duration, phrase duration, and number of syllables in a phrase did not differ significantly between deafened and intact birds for all three categorized types (Fig. 4a–c). In
addition, although it has previously been reported that the syllable repetition rate is reduced in socially isolated canaries22, in the present study no significant difference was observed
in the syllable repetition rates between deafened and intact birds, when comparing recurrent, extinct, and added phrase types separately (Fig. 4d). Taken together these results suggest that,
although the total number of acquired unique syllables in crystallized songs was decreased by auditory deprivation (Fig. 2d), the frequency of occurrence and temporal features of the three
categorized types of syllable phrasings were intrinsically regulated. AUDITORY-INDEPENDENT SONG SYNTAX IN THE CANARY We next examined the effect of auditory deprivation on song syntax in the
canary. Variations in song phrase transitions indicated a decreasing trend in deafened birds compared with intact birds, as a reduction of the total number of unique syllables was observed
following auditory deprivation (Fig. 5a). Although the phrase repertoires were not randomly generated, the probability of transition among phrases, even phrases composed of recurrent
syllables, was highly variable in both intact and deafened canaries (Fig. 5a). In addition, the probability of transition among phrases changed between the 1st and 2nd years. These results
indicate that phrase ordering was audition-independently regulated and annually reorganized. For quantitative analyses of song syntax phenotypes in the canary, we calculated consistency,
linearity, and entropy of syllable transitions in songs. No significant differences were found in the examined phenotypes of song syntax between intact and deafened birds and between the 1st
and 2nd years’ crystallized songs (Fig. 5b). This result indicates an auditory-independent recurrence of species-specific song syntax in the canary, which is similar to that observed in the
regulation of acoustic and temporal features of syllables and phrases. DISCUSSION Previous studies indicate that the canary develops a typical, species-specific song structure, which is
strictly constrained by an innate predisposition regardless of song tutoring conditions (_e.g_., isolation or artificially synthesized song)18,23,24. In the present study, we used
early-deafened canaries and showed the intrinsic, auditory-independent repeatability in the generation of an individually unique, species-specific song structure. The Waterslager canary
strain has been previously reported to show impaired high-frequency sound hearing associated with inherited reduction of hair cells25,26,27 and a lower spectral distribution of energy in the
vocalization, compared with other canary strains28. Although the elevated thresholds at high frequencies may lead to differences in song development, compared with the wild populations or
with other species, auditory deprivation clearly affected song acoustics in this strain (Fig. 2b,c)13,18. These facts indicate that Waterslager canaries use auditory information for song
development, thus contrasting with other songbird species that develop normal songs and song repertoires without appropriate auditory experiences29,30. The reduction of unique syllable types
caused by hearing loss was similar to that observed in canaries of the same strain raised in high noise environments, until deafened13. However, those canaries produced more variable
syllables than the subjects of the present study14, suggesting that the pre-deafening auditory experience might influence the generation of syllable variability after auditory deprivation.
In the present study, deafened canaries annually developed species-specific songs composing of acoustically consistent recurrent syllables. In addition, syntactical phrase ordering was
audition-independently regulated and annually reorganized. During annual song development, the phrase repertoires were reconstructed by replacing extinct syllables with newly added ones,
with no auditory input. This suggests that a predisposed innate program is crucial for the regulation of the annual syllable maintenance and replacement, which may comprise the behavioral
basis for open-ended learning. The canary annually repeats song crystallization and deterioration during adulthood, which is a characteristic of open-ended vocal learning. After song
deterioration with the collapse of acquired syllable acoustics and sequential features, the song is re-crystallized maintaining recurrent syllables, expressed in the previous crystallized
songs. Song acquisition proceeds by trial-and-error sensorimotor learning to approximate a tutored song31,32,33. Therefore, such recurrence of acoustically and temporally conserved syllables
would be feasible through auditory experiences, both for memorizing the song template and for monitoring the bird’s own vocal outputs. However, our study using early-deafened canaries
contradicts this perspective by showing an audition-independent production of recurrent song syllables. This result indicates that, rather than auditory instruction, the generation and
maintenance of individual uniqueness in canary song depends on predisposed genetic or other epigenetic programs. Although the proprioceptive feedback from the syrinx and/or respiratory
organs may be a contributing factor to guide song development in a deafened condition7,12,34, production of consistent acoustic features of syllables can only be achieved by precise
regulation of complex high-dimensional parameters, coordinating multiple syrinx muscles35. Therefore, rather than random, strict neuronal connections should be recurrently reconstructed with
reference to intrinsic information during annual song development. A large fraction of the synaptic connections between the HVC and the robust nucleus of the arcopallium RA neurons is
replaced each year from spring to fall in adult canaries36,37. However, a subpopulation of these connections and most projections from the HVC to Area X could be persistently retained
through an annual cycle of song development38,39,40. The persistent connections of HVC neurons projecting into the RA and Area X might contribute to the reproduction of consistent acoustic
and temporal features of syllables without auditory input. In addition, the replacement of new HVC neurons, which is regulated by hormones, singing, photoperiod, and neuronal activity, could
be a crucial modulator of syllable addition and loss even under auditory deprivation41. Further research is needed to investigate how such internal and external factors influence the neural
mechanisms for genetic-driven restoration of complex vocalizations. One of our motivations for this study was to compare the song of early-deafened canaries to our previous results from a
closed-ended learner, the zebra finch, which was deafened at a similar early developmental stage42. Although there are accumulated observations of song phenotypes in the oscine songbird
species after deafening, the timing of song crystallization/stabilization without auditory input has not been fully investigated. Early-deafened zebra finches show an obvious delay of song
crystallization, which takes 2- to 3-folds longer time compared with intact zebra finches42, thus indicating an auditory-dependent, active regulation of song crystallization. This
auditory-dependent regulation of song structures is also observed in the Bengalese finch, which is also a closed-ended learner43,44. However, in the canary, the auditory deprivation did not
affect the seasonal timing of song crystallization. These results clearly demonstrate that the auditory contribution to crystallization of song structures differs across species. Further
comparative experiments are needed to evaluate how the auditory experience directly affects the regulation of song crystallization and plasticity. Auditory experience enhances an individual
uniqueness to acquire a large number of unique and sexy syllable types, hence successfully attracting females45. However, individual variability in song generation is partly intrinsically
prepositioned in the canary. The degree of expression of auditory-independent individual variability in song structure was also different between the zebra finch and the canary. Deafened
zebra finches show a wider range of individual differences in both syllable acoustics and the sequential structures of their crystallized songs compared with that of intact birds42. This
individual diversification increases among different families. In contrast, deafened canaries mainly exhibited individual uniqueness at syllable acoustics, not at syllable phrasing and
transition sequence. Unlike zebra finches, auditory deprivation did not affect individual variation in the canary. This species-specific constraint for expression of individual variation
would be reflected by the history of selective breeding of canaries, on the basis of the song phenotype. Although no selective breeding has been performed in the zebra finch, the canary has
a long history spanning over 500 years of selection based on their song features, especially regarding frequency modulation of syllables and song length19,46. Therefore, the degree of
genetic variation among individuals should be different between the two species, that is, much less variability of genetic diversity in the canary than the zebra finch. This limited genetic
variation caused by inbreeding may be related to the intrinsic origin of the strict constraint in species-specific song structure, suggesting a genetic reduction of the plasticity of
individual variability in song learning. METHODS ANIMALS We used an inbred canary strain, the Belgian Waterslager, housed at the Rockefeller University Field Research Center. The birds were
raised in individual breeding cages with their parents and siblings until the 27th to the 33rd phd. By fledging, the sex of birds was checked by PCR as previously reported47. We set eight
male juveniles into two groups: auditory-intact (n = 3) and deafened (n = 5). All subjects within each group were housed together with other auditory-intact birds. The photoperiod
corresponded to the annual light cycle of the New York State, USA. Food and water were available _ad libitum_ throughout the year. All animal experiments were performed in accordance with a
protocol approved by the Rockefeller University Institutional Animal Care and Use Committee. DEAFENING The deafening operation was performed as previously described42. Five males were
deafened by bilateral cochlear extirpation at 27–34 phd (average = 29.8 phd). The birds were anesthetized with Nembutal administered by intraperitoneal injection. After fixing the head in a
custom-made stereotaxic apparatus with ear bars, a small window was opened through the neck muscle and the skull near the end of elastic extension of hyoid bone. A small hole was then made
in the cochlear dome. The cochlea was pulled out with a fine hooked wire. The removed cochleae were confirmed by visual inspection under a dissecting microscope. After cochlear removal, a
recovery period on a heat pad was set over night, after which the birds were put back into the cage. SONG RECORDING AND ANALYSIS After each bird was individually housed in a sound-proof
chamber, undirected singing was recorded once every 1–2 months using Sound Analysis Pro software, version 1.0448. Crystallized songs within a day were randomly selected. Analysis of syllable
acoustic structure was performed using the Avisoft Saslab (Avisoft Bioacoustics). Low and high frequency noise <0.5 and >19 kHz, respectively, were filtered from the recordings, and
calls were eliminated from the dataset. A song was defined as the continuous production of syllables (phrases), followed by at least 200 ms of silence. A phrase was defined as a cluster of
repetitive syllables (Fig. 1a). Analysis was performed on an average of 22.6 songs from several days per season. Syllables, the smallest unit of the song, were segmented using the Avisoft
Saslab software. The separated syllables were manually double-checked for the precision of syllable separation. To measure the complexity of song syntax as a sequence of phase transition,
consistency and linearity of phrase sequences, and its entropy of a first-order, Markov models were calculated as previously described49,50. On average, 2,576 syllables and 152.8 phrase
transitions per bird were used for the analyses. For representation of phrase transition diagrams (Fig. 5a), first, the number of phrase transition types was determined and divided by the
total number of all transitions to obtain a probability ranging from 0 to1 for each transition type. These probabilities were then graphed in a sequence transition diagram, where thicker and
darker-colored arrows indicated more frequently produced phrase transitional probabilities51. STATISTICAL ANALYSES To compare the number of syllables or phrases and the sequence of phrase
transitions between intact and deafened birds and between the two years (the 1st year versus the 2nd year), the mean values were compared using ANOVA followed by post hoc tests with Holm’s
correction. All tests were two tailed, and significance level was set at _P_ < 0.05 for all comparisons. PCA of the five syllable measurements, (1) syllable duration, (2) inter-syllable
interval, (3) fundamental frequency, (4) bandwidth, and (5) entropy, was performed using the prcomp package in the statistical software R (http://www.r-project.org/; R Core Team, 2013; R
Foundation for Statistical Computing), to investigate the difference of phrase types between the intact and deafened birds. Sampled from each bird were 8–15 phrase types (20 syllables per
phrase were used for analysis). DATA AVAILABILITY The authors confirm that all of the data underlying the reported findings are included in the article. All raw data are available from the
authors upon request. REFERENCES * Doupe, A. J. & Kuhl, P. K. Birdsong and human speech: common themes and mechanisms. _Annu Rev Neurosci_ 22, 567–631,
https://doi.org/10.1146/annurev.neuro.22.1.567 (1999). Article PubMed CAS Google Scholar * Marler, P. & Slabbekoorn, H. _Nature’s Music: The Science of Birdsong_. (Elsevier Academic
Press, 2004). * Marler, P. & Tamura, M. Culturally transmitted patterns of vocal behavior in sparrows. _Science_ 146, 1483–1486 (1964). Article ADS PubMed CAS Google Scholar * Burt,
J. M. & Beecher, M. D. The social interaction role of song in song sparrows: implications for signal design. _Comparative Cognition and Behaviour Reviews_ 3, 86–98 (2008). Google
Scholar * Miller, D. B. Long-term recognition of father’s song by female zebra finches. _Nature_ 280, 389–391 (1979). Article ADS Google Scholar * Baker, M. C. The behavioral response of
female Nuttall’s White-crowned Sparrows to male song of natal and alien dialects. _Behavioral Ecology and Sociobiology_ 12, 309–315 (1983). Article Google Scholar * Konishi, M. The role
of auditory feedback in the control of vocalization in the white-crowned saprrow. _Zeitschrift fur Tierpsychologie_ 22, 770–783 (1965). PubMed CAS Google Scholar * Marler, P. Three models
of song learning: evidence from behavior. _Journal of neurobiology_ 33, 501–516 (1997). Article PubMed CAS Google Scholar * Price, P. Developmental Deterrninants Of Structure in Zebra
Finch Song. _Jounral of Comprative and Physiological Psychology_ 93, 260–277 (1979). Article Google Scholar * Marler, P. & Sherman, V. Song structure without auditory feedback:
emendations of the auditory template hypothesis. _The Journal of Neuroscience_ 3, 517–531 (1983). Article PubMed CAS Google Scholar * Konishi, M. Effects of deafening on song development
in American robins and black-headed grobeaks. _Zeitschrift fur Tierpsychologie_ 22, 584–599 (1965). PubMed CAS Google Scholar * Konishi, M. Effects of deafening on song development in
two species of Juncos. _The conder_ 66, 85–101 (1964). Article Google Scholar * Marler, P. & Waser, M. S. Role of auditory feedback in canary song development. _Journal of comparative
and physiological psychology_ 91, 8–16 (1977). Article PubMed CAS Google Scholar * Güttinger, H. R. Self‐differentiation of Song Organization Rules by Deaf Canaries. _Ethology_ 56,
323–340 (1981). Google Scholar * Nottebohm, F., Nottebohm, M. E. & Crane, L. Developmental and seasonal changes in canary song and their relation to changes in the anatomy of
song-control nuclei. _Behavioral and Neural Biology_ 46, 445–471, https://doi.org/10.1016/S0163-1047(86)90485-1 (1986). Article PubMed CAS Google Scholar * Poulsen, H. Song learning in
the Domestic Canary. _Zeitschrift für Tierpsychologie_ 16, 173–178, https://doi.org/10.1111/j.1439-0310.1959.tb02052.x (1959). Article Google Scholar * Güttinger, H. R., Wolffgramm, J.
& Thimm, F. The Relationship Between Species Specific Song Programs and Individual Learning in Songbirds. _Behaviour_ 65, 241–261, https://doi.org/10.1163/156853978X00620 (1978). Article
Google Scholar * Waser, M. & Marler, P. Song Learning in Canaries. _Journal of Comparative and Physiological Psychology Monograph_ 91, 1–7 (1977). Article Google Scholar *
Güttinger, H. R. Consequences of domestication on the song structures in the canary. _Behaviour_ 94, 254–278 (1985). Article Google Scholar * Voigt, C. & Leitner, S. Seasonality in
song behaviour revisited: seasonal and annual variants and invariants in the song of the domesticated canary (Serinus canaria). _Hormones and behavior_ 54, 373–378,
https://doi.org/10.1016/j.yhbeh.2008.05.001 (2008). Article PubMed CAS Google Scholar * Leitner, S., Voigt, C. & Gahr, M. Seasonal changes in the song pattern of the non-domesticated
island canary (Serinus canaria) a field study. _Behaviour_ 138, 885–904 (2001). Article Google Scholar * Leitner, S. & Catchpole, C. K. Song and brain development in canaries raised
under different conditions of acoustic and social isolation over two years. _Developmental neurobiology_ 67, 1478–1487, https://doi.org/10.1002/dneu.20521 (2007). Article PubMed Google
Scholar * Gardner, T. J., Naef, F. & Nottebohm, F. Freedom and rules: the acquisition and reprogramming of a bird’s learned song. _Science_ 308, 1046–1049,
https://doi.org/10.1126/science.1108214 (2005). Article ADS PubMed CAS Google Scholar * Nottebohm, F., Stokes, T. M. & Leonard, C. M. Central control of song in the canary, Serinus
canarius. _The Journal of comparative neurology_ 165, 457–486, https://doi.org/10.1002/cne.901650405 (1976). Article PubMed CAS Google Scholar * Okanoya, K. & Dooling, R. J. Colony
differences in auditory thresholds in the canary (Serinus canarius). _The Journal of the Acoustical Society of America_ 78, 1170–1176, https://doi.org/10.1121/1.392885 (1985). Article ADS
PubMed CAS Google Scholar * Wright, T. F., Brittan-Powell, E. F., Dooling, R. J. & Mundinger, P. C. Sex-linked inheritance of hearing and song in the Belgian Waterslager canary.
_Proceedings. Biological sciences/The Royal Society_ 271(Suppl 6), S409–412, https://doi.org/10.1098/rsbl.2004.0204 (2004). Article Google Scholar * Gleich, O., Dooling, R. J. &
Manley, G. A. Inner-ear abnormalities and their functional consequences in Belgian Waterslager canaries (Serinus canarius). _Hearing research_ 79, 123–136 (1994). Article PubMed CAS
Google Scholar * Okanoya, K., Dooling, R. J. & Downing, J. D. Hearing and vocalizations in hybrid Waterslager-Roller canaries (Serinus canarius). _Hearing research_ 46, 271–275 (1990).
Article PubMed CAS Google Scholar * Kroodsma, D. E., Houlihan, P. W., Fallon, P. A. & Wells, J. A. Song development by grey catbirds. _Animal behaviour_ 54, 457–464 (1997). Article
PubMed CAS Google Scholar * Leitner, S., Nicholson, J., Leisler, B., DeVoogd, T. J. & Catchpole, C. K. Song and the song control pathway in the brain can develop independently of
exposure to song in the sedge warbler. _Proceedings. Biological sciences/The Royal Society_ 269, 2519–2524, https://doi.org/10.1098/rspb.2002.2172 (2002). Article Google Scholar *
Andalman, A. S. & Fee, M. S. A basal ganglia-forebrain circuit in the songbird biases motor output to avoid vocal errors. _Proceedings of the National Academy of Sciences of the United
States of America_ 106, 12518–12523, https://doi.org/10.1073/pnas.0903214106 (2009). Article ADS PubMed PubMed Central Google Scholar * Fee, M. S. & Goldberg, J. H. A hypothesis for
basal ganglia-dependent reinforcement learning in the songbird. _Neuroscience_ 198, 152–170, https://doi.org/10.1016/j.neuroscience.2011.09.069 (2011). Article PubMed PubMed Central CAS
Google Scholar * Charlesworth, J. D., Warren, T. L. & Brainard, M. S. Covert skill learning in a cortical-basal ganglia circuit. _Nature_ 486, 251–255,
https://doi.org/10.1038/nature11078 (2012). Article ADS PubMed PubMed Central CAS Google Scholar * Suthers, R. A., Goller, F. & Wild, J. M. Somatosensory feedback modulates the
respiratory motor program of crystallized birdsong. _Proceedings of the National Academy of Sciences of the United States of America_ 99, 5680–5685, https://doi.org/10.1073/pnas.042103199
(2002). Article ADS PubMed PubMed Central CAS Google Scholar * Suthers, R. A. & Zollinger, S. A. Producing song: the vocal apparatus. _Annals of the New York Academy of Sciences_
1016, 109–129 (2004). Article ADS PubMed Google Scholar * Kirn, J. R., Alvarez-Buylla, A. & Nottebohm, F. Production and survival of projection neurons in a forebrain vocal center of
adult male canaries. _The Journal of neuroscience: the official journal of the Society for Neuroscience_ 11, 1756–1762 (1991). Article CAS Google Scholar * Nottebohm, F., O’Loughlin, B.,
Gould, K., Yohay, K. & Alvarez-Buylla, A. The life span of new neurons in a song control nucleus of the adult canary brain depends on time of year when these cells are born.
_Proceedings of the National Academy of Sciences of the United States of America_ 91, 7849–7853 (1994). Article ADS PubMed PubMed Central CAS Google Scholar * Kirn, J. R. &
Nottebohm, F. Direct evidence for loss and replacement of projection neurons in adult canary brain. _The Journal of neuroscience:the official journal of the Society for Neuroscience_ 13,
1654–1663 (1993). Article PubMed CAS Google Scholar * Gahr, M. Delineation of a brain nucleus: comparisons of cytochemical, hodological, and cytoarchitectural views of the song control
nucleus HVc of the adult canary. _Journal of Comparative Neurology_ 294, 30–36 (1990). Article PubMed CAS Google Scholar * Alvarez-Buylla, A., Theelen, M. & Nottebohm, F. Birth of
projection neurons in the higher vocal center of the canary forebrain before, during, and after song learning. _Proceedings of the National Academy of Sciences of the United States of
America_ 85, 8722–8726 (1988). Article ADS PubMed PubMed Central CAS Google Scholar * Balthazart, J. & Ball, G. F. Endocrine and social regulation of adult neurogenesis in
songbirds. _Frontiers in neuroendocrinology_ 41, 3–22, https://doi.org/10.1016/j.yfrne.2016.03.003 (2016). Article PubMed CAS Google Scholar * Mori, C. & Wada, K.
Audition-Independent Vocal Crystallization Associated with Intrinsic Developmental Gene Expression Dynamics. _Journal of Neuroscience_ 35, 878–889,
https://doi.org/10.1523/jneurosci.1804-14.2015 (2015). Article PubMed Google Scholar * Okanoya, K. & Yamaguchi, A. Adult Bengalese finches (Lonchura striata var. domestica) require
real-time auditory feedback to produce normal song syntax. _Journal of neurobiology_ 33, 343–356 (1997). Article PubMed CAS Google Scholar * Woolley, S. M. N. & Rubel, E. W.
Bengalese Finches Lonchura Striata Domestica Depend upon Auditory Feedback for the Maintenance of Adult Song. _The Journal of Neuroscience_ 17, 6380–6390 (1997). Article PubMed CAS Google
Scholar * Vallet, E. & Kreutzer, M. Female canaries are sexually responsive to special song phrases. _Animal behaviour_ 49, 1603–1610, https://doi.org/10.1016/0003-3472(95)90082-9
(1995). Article Google Scholar * Mundinger, P. Behaviour genetic analysis of selective song learning in three inbred canary strains. _Behaviour_ 147, 705–723,
https://doi.org/10.1163/000579510x489903 (2010). Article Google Scholar * Wada, K. _et al_. A molecular neuroethological approach for identifying and characterizing a cascade of
behaviorally regulated genes. _Proceedings of the National Academy of Sciences of the United States of America_ 103, 15212–15217, https://doi.org/10.1073/pnas.0607098103 (2006). Article ADS
PubMed PubMed Central CAS Google Scholar * Tchernichovski, O., Mitra, P. P., Lints, T. & Nottebohm, F. Dynamics of the vocal imitation process: how a zebra finch learns its song.
_Science_ 291, 2564–2569, https://doi.org/10.1126/science.1058522 (2001). Article ADS PubMed CAS Google Scholar * Scharff, C. & Nottebohm, F. A comparative study of the behavioral
deficits following lesions of various parts of the zebra finch song system: implications for vocal learning. _The Journal of neuroscience: the official journal of the Society for
Neuroscience_ 11, 2896–2913 (1991). Article CAS Google Scholar * Lemon, R. E., Dobson, C. W. & Clifton, P. G. Songs of American Redstarts (Setophaga ruticilla): Sequencing Rules and
their Relationships to Repertoire Size. _Ethology_ 93, 198–210, https://doi.org/10.1111/j.1439-0310.1993.tb00989.x (1993). Article Google Scholar * Horita, H., Wada, K. & Jarvis, E. D.
Early onset of deafening-induced song deterioration and differential requirements of the pallial-basal ganglia vocal pathway. _The European journal of neuroscience_ 28, 2519–2532,
https://doi.org/10.1111/j.1460-9568.2008.06535.x (2008). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS We thank Dr. Fernando Nottebohm for providing
the canaries; Sharon Sepe and Helen Ecklund for bird keeping at The Rockefeller University Field Research Center; Akiko Ueno and Raimu Imai for helping initial song recording and analysis;
and M. Sanchez, A.N. Chinweike, and S. Hayase for critical reading. The work was supported by a Grant-in-Aid for JSPS Fellows 11J07071 to C.M. and Takeda Science Foundation, Inamori
Foundation, JSPS KAKENHI (16H01261, 17H05932, 18H02520, and 17K19629), and by a MEXT Grant-in-Aid for Scientific Research on Innovative Areas #4903 (Evolinguistics; JP17H06383) to K.W.
AUTHOR INFORMATION Author notes * Chihiro Mori Present address: Department of Life Sciences, Graduate School of Arts and Sciences, The University of Tokyo, Tokyo, Japan AUTHORS AND
AFFILIATIONS * Graduate School of Life Science, Hokkaido University, Sapporo, Hokkaido, Japan Chihiro Mori & Kazuhiro Wada * Department of Biological Sciences, Hokkaido University,
Sapporo, Hokkaido, Japan Kazuhiro Wada * Faculty of Science, Hokkaido University, Sapporo, Hokkaido, Japan Kazuhiro Wada * Department of Psychology, Colgate University, Hamilton, New York,
USA Wan-chun Liu Authors * Chihiro Mori View author publications You can also search for this author inPubMed Google Scholar * Wan-chun Liu View author publications You can also search for
this author inPubMed Google Scholar * Kazuhiro Wada View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS C.M. and K.W. designed the research.
C.M. performed the experiments. W.L. helped with song recording. C.M. and K.W. performed the analysis and wrote the paper. CORRESPONDING AUTHOR Correspondence to Kazuhiro Wada. ETHICS
DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE: Springer Nature remains neutral with regard to jurisdictional
claims in published maps and institutional affiliations. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which
permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to
the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless
indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or
exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints
and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Mori, C., Liu, Wc. & Wada, K. Recurrent development of song idiosyncrasy without auditory inputs in the canary, an open-ended vocal
learner. _Sci Rep_ 8, 8732 (2018). https://doi.org/10.1038/s41598-018-27046-4 Download citation * Received: 19 June 2017 * Accepted: 24 May 2018 * Published: 07 June 2018 * DOI:
https://doi.org/10.1038/s41598-018-27046-4 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative