Play all audios:
ABSTRACT Cell survival is facilitated by the maintenance of mitochondrial membrane potential (MMP). B cell activating factor (BAFF) plays a role in survival, differentiation, and maturation
of B cells. In the present study, we examined whether BAFF could attenuate oxidative stress-induced B cell death by the regulation of MMP collapse via spleen tyrosine kinase (Syk) activation
using WiL2-NS human B lymphoblast cells. BAFF binds to receptors on WiL2-NS cells. When the cells were incubated in serum-deprived conditions with 1% fetal bovine serum (FBS), BAFF reduced
the percentage of dead cells as determined through trypan blue staining and caspase 3 activity. BAFF also inhibited MMP collapse with 1% FBS, as indicated by a decrease in the number of
cells with high-red fluorescence of MitoProbe™ JC-1 reagent or a decrease in the percentage of DiOC6-stained cells. Reactive oxygen species (ROS) production was reduced by incubation with
BAFF in the presence of 10% or 1% FBS. BAFF inhibited MMP collapse, cell growth retardation, dead cell formation, and caspase 3 activation caused by treatment with H2O2. Syk phosphorylation
on tyrosine (Y) 525/526 was increased in cells incubated with 1% FBS in the presence of BAFF than cells incubated with 1% FBS or BAFF alone. BAY61-3606, a Syk inhibitor reduced the effect of
BAFF on MMP collapse, caspase 3 activation, cell growth retardation, and dead cell formation. Together, these data demonstrate that BAFF might attenuate oxidative stress-induced B cell
death and growth retardation by the maintenance of MMP through Syk activation by Y525/526 phosphorylation. Therefore, BAFF and Syk might be therapeutic targets in the pathogenesis of B
cell-associated diseases such as autoimmune disease. SIMILAR CONTENT BEING VIEWED BY OTHERS IMMUNE STRESS SUPPRESSES INNATE IMMUNE SIGNALING IN PRELEUKEMIC PRECURSOR B-CELLS TO PROVOKE
LEUKEMIA IN PREDISPOSED MICE Article Open access 24 August 2023 EXTRACELLULAR VESICLES FROM TYPE-2 MACROPHAGES INCREASE THE SURVIVAL OF CHRONIC LYMPHOCYTIC LEUKEMIA CELLS EX VIVO Article
Open access 25 June 2024 THIOREDOXIN IS A METABOLIC RHEOSTAT CONTROLLING REGULATORY B CELLS Article Open access 29 March 2024 INTRODUCTION B cell activating factor (BAFF), also known as
TALL-1, BLyS, THANK, and TNFSF13B, belongs to the tumor necrosis factor (TNF) superfamily1. BAFF is expressed in both immune cells such as dendritic cells, macrophages, and T cells1 and
non-immune cells such as alveolar-associated cells2, fibroblast-like synoviocytes3, and various epithelial cells4. BAFF naturally exists as a transmembrane protein and is solubilized by
proteolytic cleavage of furin convertase between R133 and A1345,6. BAFF transmits signals by binding to BAFF receptors, BAFF-R, transmembrane activator and calcium-modulator and cytophilin
ligand interactor (TACI), and B cell maturation antigen (BCMA), with different binding affinities. Weak BAFF stimuli cause a deficiency of mature B cells7,8. In contrast, excessive systemic
humoral immune responses by elevated BAFF levels lead to the accumulation of antigen–antibody immune complexes, which in turn lead to inflammatory responses and aggravate autoimmune diseases
such as rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), and type 1 diabetes1,9,10. Inflammation is a general symptom of autoimmune disease and induces large quantities of
reactive oxygen species (ROS) in disease sites11,12,13. Highly reactive unpaired electrons in ROS can cause tissue damage, leading to apoptosis or necrosis14. In the process of cell death,
the electron transport chain in the mitochondria is disrupted15, resulting in the collapse of the mitochondria membrane potential (MMP), and the hydrogen ion concentration gradient between
the mitochondrial intermembrane space and the matrix16. The maintenance of MMP is crucial for ATP production17 and cell survival15. Therefore, the collapse of MMP is regarded as a
pro-apoptosis signal18. BAFF also reportedly up-regulates MMP through protein kinase B (AKT) and PKC-β in mouse B cells19. Meanwhile, another study found that BAFF expression was increased
by ROS in mouse macrophages20. However, little is known about the effects of BAFF on ROS production in human B cells. Spleen tyrosine kinase (Syk) is a major signal transmitter that
contributes to B cell survival and differentiation21,22. Syk consists of many tyrosine (Y) residues that can be phosphorylated, and the phosphorylation of Y323, Y352, and Y525/526 leads to
various pathways9,23,24,25. Among the phosphorylation sites in Syk26, Y525/526 is in a kinase domain that is important for kinase activity27. Syk is activated by ROS28,29,30 in chicken B
cells and BAFF in mouse B cells31. However, little is known about the effects of BAFF on MMP by ROS production via Syk in human B cells. The role of Syk on cell fate varies depending on the
cell type and cellular conditions. Oxidative stress-induced Syk activation activates different pathways, such as pro-apoptotic and survival pathways. Therefore, a balance of pro-apoptotic
and survival pathways are important for determining the fate of cells exposed to oxidative stress13. Syk expression protects cells from apoptosis induced by oxidative or genotoxic stress in
MCF7 and MDA-MB-231 breast cancer cells and DG75 B-lymphoma cells32. Syk-deficient B cells can survive in the periphery for an extended period of time33. Syk inhibition is a promising
therapeutic strategy to disrupt pro-survival microenvironment signaling in chronic lymphocytic leukemia34. Syk is also involved in the induction of G2/M arrest, which protects cells from
apoptosis. Syk-dependent PLC-gamma2 activation was required for acceleration towards apoptosis following oxidative stress13. These reports from different groups yielded contradictory
results, so it is necessary to clarify the connection between BAFF signaling and Syk phosphorylation in B cells. In this study, we investigated whether BAFF could ameliorate B cell survival
through the inhibition of ROS production, MMP collapse, and Syk activation. To do this, we incubated WiL2-NS human B lymphoblast cells isolated from the spleen of a Caucasian male (American
Type Culture Collection) in serum-deprived conditions (1% FBS). Our results suggest that Syk is a key regulator for the maintenance of MMP by BAFF in human B cells. MATERIALS AND METHODS
REAGENTS Recombinant human BAFF, biotinylated anti-human BAFF-R and TACI antibodies was purchased from R&D Systems (Minneapolis, MN, USA). Human BAFF-murine CD8 (BAFF-muCD8),
biotinylated fusion proteins, were purchased from Ancell (Bayport, MN, USA). BAY61-3606, Syk inhibitor, was purchased from Adooq bioscience (Irvine, CA, USA). Catalase, dimethyl sulfoxide
(DMSO), _N_-acetyl-L-cysteine (NAC), hydrogen peroxide (H2O2) and 3,3′-dihexyloxacarbocyanine iodide (DiOC6) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolocarbocyanine iodide (JC-1) was obtained from Invitrogen (Eugene, Oregon, USA). 2′,7′–dichlorofluorescin diacetate (DCF-DA) was
purchased from Molecular Probe (Eugene, Oregon, USA). Ac-DEVD-pNA, caspase 3 substrate, was obtained from Santa Cruz Biotechnology (Santa cruz, CA, USA). Antibodies to Syk and phospho-Syk
(Y525/526) were purchased from Cell Signaling Technology (Berverly, MA, USA). Except indicated, all chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA). CELL CULTURES WiL2-NS
(ATCC® CRL-8155™), a human B lymphoblast cells were provided from the Korea Research Institute of Bioscience and Biotechnology (KRIBB) cell bank (Daejeon, Korea). Cells were incubated in
RPMI 1640 medium (GIBCO, Grand Island, NY, USA) with 10% heat-inactivated fetal bovine serum (FBS) (GIBCO, Grand Island, NY, USA), 2 mM L-glutamine and 100 units/ml of
penicillin/streptomycin (GIBCO, Grand Island, NY, USA) at 37 °C humidified incubator under the 5% CO2 condition35. MEASUREMENT OF BAFF-BAFF RECEPTOR INTERACTIONS BAFF binding on BAFF
receptors were determined by the method previously described36. Briefly, WIL2-NS cells were suspended in 100 μl of Hank’s Balanced Salt Solution (HBSS) complemented with 2% bovine calf serum
(BCS). Then, cells were incubated with 10 μg/ml of a human BAFF-murine CD8 (BAFF-muCD8) biotinylated fusion protein on ice for 30 min. After washing cells with 2% BCS-containing HBSS twice,
cells were incubated with phycoerythrin (PE)-conjugated streptavidin (BD pharmingen, SanJose, CA, USA) for 20 min on ice. After washing cells with 2% BCS-containing HBSS twice, fluorescence
intensity of 10,000 cells was analyzed by FACSCalibur™ (Becton Dickinson, San Joes, CA, USA). MEASUREMENT OF REACTIVE OXYGEN SPECIES Intracellular ROS level was measured by incubating cells
with or without 10 μM DCF-DA at 37 °C for 30 min. Flourescence intensity of 10,000 cells was analyzed by FACSCalibur™ (Becton Dickinson, San Joes, CA, USA)35. MEASUREMENT OF MITOCHONDRIAL
MEMBRANE POTENTIAL To measure mitochondrial membrane potential (MMP), cells were stained with 2.5 μg/ml JC-1 or DiOC6 for 10 min at 37 °C. Then, cells stained with JC-1 were observed by 485
nm filter of fluorescence microscope and five areas were pictured to count number of high red fluorescent (J-aggregate) live cells. Dead or dying cells exhibit yellow or green fluorescence
(J-monomer) with collapsed mitochondrial potential, respectively. Data were represented by the percentage of high red-fluorescence cells37. In addition, cells stained with DiOC6 were
analyzed by FACSCalibur™ (Becton Dickinson, San Joes, CA, USA). Cells with MMP collapse showed a decrease in DiOC6 fluorescence intensity. MEASUREMENT OF CASPASE 3 ACTIVITY According to
previous report37, 1 × 107 cells were harvested and lysed in 100 μl lysis buffer containing 50 mM 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid (HEPES, pH 7.4), 5 mM
(3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS), and 5 mM dithiothreitol (DTT). for 20 min on ice. Then, cell lysates were centrifuged by 16,000 × _g_ for 10 min at 4 °C.
After centrifugation, supernatants were transferred into new tube. Each 5 μl supernatant sample was incubated with 200 μM of caspase 3 substrate (Ac-DEVD-pNA) in assay buffer containing 20
mM HEPES (pH 7.4), 0.1% CHAPS, 5 mM DTT, and 2 mM ethylenediaminetetraacetic acid (EDTA). Total incubation volume was 100 μl per each well in 96 well plate, After incubation for 2 h, optical
density was measured by ELISA reader (Molecular Devices, Sunnyvale, CA) at 405 nm. Each sample was normalized by protein concentration. TRYPAN BLUE EXCLUSION ASSAY Diluted cell suspension
was mixed with equal volume of 0.4% trypan blue in PBS. Dying or dead cells were stained with blue color and viable cells were unstained. Each cell was counted by using hemocytometer under
light microscope (Olympus Korea Co., Ltd, Seoul, Rep. of Korea)35. WESTERN BLOT ANALYSIS As reported previously35, cellular proteins were extracted by 0.5% Nonidet P-40 lysis buffer
containing 20 mM Tris-HCl (pH 8.2), 150 mM NaCl, protease inhibitor (2 μg/ml aprotinin, 2 μg/ml pepstatin, 1 μg/ml leupeptin, 1 mM phenylmethylsulfonyl fluoride) and phosphatase inhibitor (1
mM sodium vanadate and 5 mM sodium fluoride). Cells were lysed for 30 min on ice and centrifuged at 13,000 rpm for 20 min at 4 °C. Protein concentrations of lysates were determined by using
SMART™ BCA protein assay kit (iNtRON, Gyeonggido, Korea). Equal amounts of cellular proteins in sodium dodecyl sulfate (SDS) sample buffer were denatured by boiling at 100 °C for 5 min.
Samples were separated according to protein size by SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis). Separated samples were transferred to nitrocellulose membrane.
Membranes were blocked with 2% skim milk in Tris buffered saline containing 0.5% Tween20. After blocking, protein expression of each sample was probed by immune-reaction using enhanced
chemiluminescence. STATISTICAL ANALYSES Experimental differences were examined separately for statistical significance using ANOVA and Students’ t-distribution. The _p_ value of < 0.05 or
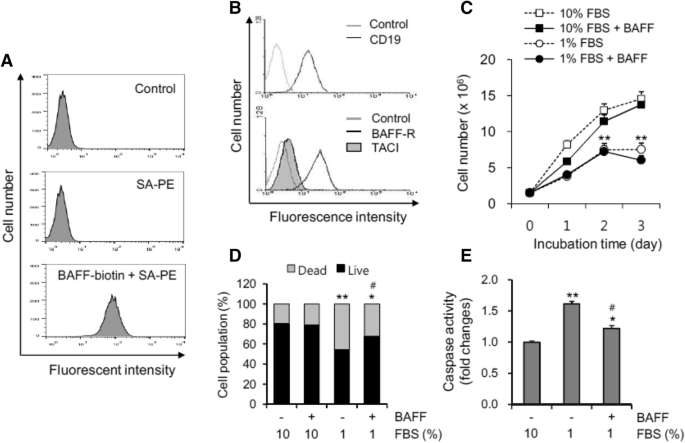
< 0.01 was considered to be significant35,37. RESULTS BAFF RESCUED CELLS FROM OXIDATIVE STRESS-INDUCED CELL DEATH To examine the effects of BAFF on a human B cell line, we determined
whether BAFF could bind to the cell surface of WiL2-NS human B cells that expressed BAFF receptors. To do this, cells were incubated with biotin-labelled BAFF or antibodies to BAFF
receptors. The results revealed a significant increase in BAFF binding on the surface of WiL2-NS cells (Fig. 1A). The expression level of BAFF-R was higher than that of TACI (Fig. 1B).
However, no BCMA expression was detected (data not shown), which suggests that BAFF might affect WiL2-NS cells via BAFF binding on the cell surface. Next, we used WiL2-NS cells to assess the
effect of BAFF on molecular and cellular changes that control B cell death or survival. BAFF increases B cell survival1,9,10 and is associated with an increase in the incidence of
autoimmune disease. Additionally, serum deprivation (SD) leads to apoptotic cell death38,39; therefore, we examined the effects of BAFF on oxidative stress-induced cell death by using 1%
rather than 10% FBS. As shown in Fig. 1C, the cell number was reduced to about half by incubation with 1% FBS compared to 10% FBS. When cells were incubated with 10% or 1% FBS-containing
medium in the presence or absence of BAFF, no changes were detected in total cell number. The number of dead cells was determined using a trypan blue staining assay. The percentage of cell
survival with 1% FBS decreased to about 50% as compared to about 80% with 10% FBS. When cells were incubated with 1% FBS-containing medium in the presence of BAFF, the percentage of cell
survival was rescued to about 70% (Fig. 1D). In 1% FBS, a caspase 3 assay revealed that cell death increased by about 1.6 times compared to that with 10% FBS. Caspase 3 activity induced by
1% FBS decreased about 1.2 times by treatment with BAFF (Fig. 1E). This suggests that BAFF binding might activate or change molecules or intracellular events to control B cell death. BAFF
ATTENUATED MMP COLLAPSE IN SERUM-DEPRIVED (SD) CONDITIONS MMP plays a role in the regulation of various cellular functions37,40, and SD induces apoptotic cell death of rat retinal ganglion
cells via mitochondrial changes38. Therefore, we determined the effects of BAFF on MMP changes using MitoProbe™ JC-1 reagent in WiL2-NS cells that were incubated with 1% FBS. The number of
live cells with high red fluorescence was determined to assess the MMP of cells incubated with 1% FBS-containing medium. The MMP decreased to about 40% as compared to 83% with 10% FBS. When
cells were incubated with 1% FBS-containing medium in the presence of BAFF, the MMP was significantly rescued to about 60% (Fig. 2A). MMP changes by 1% FBS were also determined by incubation
with DiOC6. As shown in Fig. 2B, the fluorescence intensity of DiOC6 was decreased by incubation with 1% FBS. The percentage of cells with low fluorescence intensity of DiOC6 increased to
about 36% as compared to 0.5% with 10% FBS. The mean fluorescence intensity (MFI) of the cell population decreased to about 50.5 as compared to 106.8 with 10% FBS. When cells were incubated
with 1% FBS-containing medium in the presence of BAFF, the percentage of cells with low fluorescence intensity of DiOC6 decreased to about 29% and the MFI of the cell population increased to
about 58.6. This suggests that BAFF could regulate B cell survival by controlling MMP via BAFF-mediated activation of molecules. BAFF REDUCED ROS PRODUCTION IN SERUM-DEPRIVED (SD)
CONDITIONS Because ROS are reportedly the main cause of MMP collapse in various cells16,37,38,40,41, we examined the changes in intracellular ROS levels by incubation with 1% FBS using
dihydrodichlorofluorescein diacetate (DCF-DA). As shown in Fig. 3A, the ROS level was significantly increased by incubation with 1% FBS as compared with 10% FBS. The MFI of the cell
population in 10% and 1% FBS was 114 and 377, respectively. When cells were incubated with 1% FBS-containing medium in the presence of BAFF, the ROS level significantly decreased, as judged
by a MFI of 311. This suggests that BAFF could regulate B cell survival by controlling MMP via BAFF binding-mediated regulation of ROS production. The increase in ROS level in the presence
of 1% FBS was reduced by treatment with catalase, which catalyzes the decomposition of hydrogen peroxide (H2O2) to water and oxygen. The MFI of the cell population in 1% FBS was 195.5, which
was reduced to 123.1 by incubation with catalase as compared to 53.2 in the control group in 10% FBS (Fig. 3B). As shown in Fig. 3C, catalase also reduced the extent of MMP collapse, which
was determined by incubation with DiOC6. The MFI of the cell population in 1% FBS was 52.9, which increased to 63.7 by incubation with catalase as compared to 103.8 in the control group (10%
FBS). In addition, the percentage of cells with low fluorescence of DiOC6 in 1% FBS was 31.4%, which decreased to 9.7% as compared to 0.2% in the control group. This suggests that cell
death might be mediated by ROS, which can cause H2O2-induced MMP collapse. BAFF INHIBITED H2O2-INDUCED MMP COLLAPSE AND B CELL DEATH To confirm the inhibitory effect of BAFF on ROS-induced
MMP collapse and cell death, WiL2-NS cells were incubated with 100 µM H2O2. MMP collapse by H2O2 treatment was assessed by staining cells with MitoProbe™ JC-1 reagent and by counting the
number of live cells with high red fluorescence. H2O2 treatment decreased the MMP to about 46% as compared to 82% in the control group. When cells were incubated with 100 µM H2O2 in the
presence of BAFF, MMP collapse was inhibited to about 60% (Fig. 4A). Caspase 3 assay results revealed that cell death in the presence of 100 µM H2O2 increased by about 1.5 times as compared
to the control group. H2O2-induced caspase 3 activity was reduced about 1.2 times by treatment with BAFF (Fig. 4B). Incubation with H2O2 reduced the total cell number by about 40% compared
to the control group, which was increased slightly in the presence of BAFF (Fig. 4C). Using trypan blue staining to assess the number of dead cells, the percentage of cell survival was
reduced to about 89% as compared to about 96% in 10% FBS. When cells were incubated with H2O2 in the presence of BAFF, the percentage of cell survival was rescued to about 95% (Fig. 4D).
This suggests that BAFF binding might activate or change molecules, or alter intracellular events to control H2O2-induced B cell death. SYK PHOSPHORYLATION AT Y525/526 WAS ACTIVATED BY BAFF
OR SERUM DEPRIVATION Because Syk can be activated by BAFF42 and plays an important role in B cell survival42, we sought to identify the tyrosine site of Syk that might be activated by BAFF
binding to WiL2-NS lymphoblast B cells. When cells were incubated with 20 ng/ml of BAFF, we observed a time-dependent increase in Syk phosphorylation at Y525/526 (Fig. 5A). However, few
changes were detected in Syk phosphorylation at Y323 or Y352. Syk phosphorylation at Y525/526 was enhanced by incubation with 1% FBS (Fig. 5B). When cells were incubated with 1% FBS in the
presence of BAY61-3606, a Syk inhibitor, we observed a decrease in Syk phosphorylation at Y525/526 (Fig. 5C). In contrast, when cells were incubated with 1% FBS-containing medium in the
presence of BAFF, Syk phosphorylation at Y525/526 was greatly increased compared to that in the control group with 1% FBS (Fig. 5D). These results indicate that BAFF provided a much stronger
signal for B cell survival via Syk phosphorylation. Therefore, Syk activation might be required for B cells to survive in an environment that induces cell death. SYK INHIBITOR ENHANCED MMP
COLLAPSE IN SERUM-DEPRIVED CONDITIONS Because Syk is localized in the mitochondrial inter-membrane space where it can regulate respiratory activity42, we tested the role of BAFF-mediated Syk
activation on MMP collapse in WiL2-NS cells using MitoProbe™ JC-1 reagent under serum-deprived conditions (1% FBS). Next, cells were incubated with 1% FBS-containing medium in the presence
of BAY61-3606 and/or BAFF. The number of live cells with high red fluorescence was counted to assess the MMP (Fig. 6A, top). The results revealed that the MMP with 1% FBS decreased to about
18% as compared to 93% with 10% FBS. Treatment with BAFF significantly rescued the MMP to about 32%, which was inhibited by BAY61-3606. However, few changes were observed by treatment with
BAY61-3606 alone (Fig. 6A, bottom). DiOC6 was also used to confirm the effect of the Syk inhibitor on MMP collapse. As shown in Fig. 6B, the fluorescence intensity of DiOC6 was decreased by
incubation with 1% FBS. The percentage of cells with low fluorescence intensity of DiOC6 was increased to about 28.3%, which was reduced to about 20.6% by incubation with BAFF as compared to
3.2% with 10% FBS (Fig. 6B, top). The MFI of the cell population with 1% FBS was 78.5, which was increased to about 88.7 by incubation with BAFF as compared to about 106.6 with 10% FBS.
Additionally, when cells were incubated with 1% FBS in the presence of BAY61-3606, the number of cells with low fluorescence intensity of DiOC6 significantly increased to about 60.7%, which
was recovered to 53.4% by incubation with BAFF. The MFI of the cell population in the presence of BAY61-3606 was decreased to about 61.1, which was also enhanced to 68.5 by incubation with
BAFF (Fig. 6B, bottom). This suggests that MMP collapse could be maintained by BAFF via Syk activation. BAFF-MEDIATED INHIBITION OF B CELL DEATH WAS ATTENUATED BY SYK INHIBITION To confirm
the role of Syk activation by BAFF in the inhibition of B cell death, WiL2-NS cells were incubated with 1% FBS-containing medium in the presence of BAY61-3606 and/or BAFF. Caspase 3 activity
was enhanced by 1% FBS about 1.8 times as compared to that with 10% FBS. Caspase 3 activity induced by 1% FBS was reduced to about 1.2 times by treatment with BAFF, which was inhibited by
BAY61-3606. Additionally, almost the same changes in caspase 3 activity were observed by treatment with BAY61-3606 alone (Fig. 7A). The total cell number was reduced to about 70% by
incubation with 1% FBS as compared to that with 10% FBS. When cells were incubated with 1% FBS-containing medium in the presence or absence of BAFF, the total cell number decreased by
incubation with 1% FBS was increased to about 90% by treatment with BAFF. However, the total cell number was highly reduced by treatment with BAY61-3606 in the presence or absence of BAFF
(Fig. 7B). Trypan blue staining was used to determine the number of dead cells; the percentage of cell survival was calculated and was found to be reduced about 92% with 1% FBS as compared
to about 98% with 10% FBS, which was inhibited by BAY61-3606. When cells were incubated with 1% FBS-containing medium in the presence of BAFF, the percentage of cell survival was rescued to
about 99%, which was also inhibited by BAY61-3606 (Fig. 7C). These results were consistent with changes in Syk phosphorylation at Y525/526 (Fig. 7D, left). When cells were incubated with 1%
FBS-containing medium, Syk phosphorylation at Y525/526 increased about 2.5-fold as compared to control with 10% FBS, which was inhibited by BAY61-3606. Syk phosphorylation at Y525/526
increased about 4.7-fold by incubation with 1% FBS in the presence of BAFF, which was also inhibited by BAY61-3606 (Fig. 7D, right). This suggests that BAFF might regulate B cell survival by
controlling MMP via oxidative stress-induced Syk activation (Fig. 7E). DISCUSSION B cells play a role in the progression of autoimmune disease by releasing inflammatory cytokines,
presenting autoantigens, producing antibodies, and ensuring interactions with T cells43. Therefore, many researchers are investigating how B cells can survive in the aggravation of
autoimmune disease. ROS are increased in most inflammatory disease sites and are highly reactive due to their unpaired electron11,12,13,14. ROS that are excessively increased in inflammatory
responses could affect cell fate by damaging various cellular components such as DNA, lipids, proteins13,14, and organelles15. For example, the electron transport chain in mitochondria can
be disrupted in the middle of cell death15. BAFF enhances B cell survival, which is indispensable for B cell maturation and the enhancement of immune responses. Excessive responses by
elevated BAFF levels lead to the accumulation of antigen–antibody immune complexes, which aggravate autoimmune diseases such as RA, SLE, and type 1 diabetes1,9,10. ROS can induce B cell
activation, which is similar to the response mediated via B cell antigen receptors. Syk is a molecule that is activated by ROS28,29. However, little is known about the regulation of B cell
survival by Syk activation-associated MMP control via ROS induction. BAFF mediates ROS control: here, we investigated whether B cell survival could be regulated via BAFF-mediated control of
ROS and MMP collapse via Syk activation using a WiL2-NS human B cell line. Our results revealed that BAFF ameliorated SD-induced B cell death by the inhibition of MMP collapse through Syk
activation. Therefore, Syk could be a key regulator for the maintenance of MMP by BAFF for human B cell survival. BAFF can bind three receptors: BAFF-R, TACI, and BCMA. Among these, BAFF-R
has binding affinity for only BAFF, and BCMA has the weakest binding affinity. TACI and BCMA also have affinity to APRIL, which has a similar structure to BAFF1. Our results revealed that
BAFF binding was significant on the cell surface of WiL2-NS human B lymphocytes. While WiL2-NS cells expressed high levels of BAFF-R and low levels of TACI, no BCMA expression was detected.
BAFF has a different binding affinity to each BAFF receptor, so it is necessary to determine how much each receptor contributes to BAFF-induced survival. Although the exact cause of
autoimmune disease is not well-understood, auto-antibodies from auto-reactive B cells that attack host cells are cited as one of the main causes44,45. To produce autoantibodies, B cells must
overcome many harmful kinds of environments such as those with a high amount of ROS or MMP collapse, which threaten cell death in disease sites. BAFF enhances B cell survival, which is
indispensable for B cell immune responses that aggravate autoimmune diseases 1,9,10. Our research, conducted in serum-deprived conditions, provides the groundwork for an experimental model
to test the effect of therapeutic candidates on BAFF-mediated autoimmune disease-like states with ROS production and MMP collapse. BAFF up-regulated MMP through AKT and PKC-β in mouse B
cells19. BAFF also activated Syk31 in B cells of different species. BAFF expression was also increased by ROS in mouse macrophages20. Therefore, it is possible for BAFF expression to be
regulated by Syk activation. Syk belongs to the Syk family of tyrosine kinases and transmit signals from B-cell receptor (BCR), T-cell receptor (TCR), and BAFF receptors for various cellular
responses26. Syk consists of many tyrosine residues that can potentially be phosphorylated. In human Syk, Y323, Y352, and Y525/526 are the most reported residues. Each of the tyrosine
phosphorylation sites leads to a different downstream pathway. Phosphorylated Y323 becomes the binding site for the E3 ubiquitin ligase CBL, which mediates Syk ubiquitylation and degradation
9,23,25. Phosphorylated Y352 is the binding site for the PI3K regulatory subunit p85α N-terminal SH2 domain. It is also the binding site for the C-terminal SH2 domain of PLC-γ24. Y525/526
is located in the kinase domain, and its phosphorylation plays a crucial role in Syk activation27. Our results indicate that SD or BAFF treatment increased Syk phosphorylation at Y525/526.
This suggests that BAFF-mediated cell survival could be dependent on Syk activation to control MMP collapse by SD. Syk also contributes to B cell survival and differentiation21,22.
Therefore, Syk activation in our study could be associated with different molecules to maintain cell survival. Syk might inhibit the activation of caspase-9 through AKT13. AKT plays an
important role in B cell survival and is activated in a Syk-dependent pathway46. Syk also protects cells from apoptosis induced by oxidative or genotoxic stress by stabilizing the mRNA for
Bcl-x(L), an antiapoptotic protein32. Syk also transduces BAFF survival signals via extracellular signal-regulated kinase (ERK) and PI3 kinase31. At the same time, the possibility that Syk
might activate cell death pathways cannot be ruled out. Syk-dependent PLCγ2 activation was required for acceleration towards apoptosis following oxidative stress13. Mule (also known as
ARF-BP1, HUWE1, Ureb1, LASU1, and HECTH9) belongs to the E5-AP C terminus (HECT) domain-containing ubiquitin ligase (E3). Mule phosphorylation by Syk induces TNF-induced c-Jun N-terminal
kinase (JNK) activation and cell death47. As such, it is necessary to define Syk-associated molecules by BAFF binding on cell surfaces in the inhibition of SD-induced MMP collapse. In
conclusion, our findings demonstrate that SD-induced ROS production causes MMP collapse through oxidative stress, which leads to cell death. Furthermore, this was attenuated by BAFF binding
on BAFF receptors via Syk activation. Together, our results suggest that Syk activation by BAFF-BAFF receptor binding could regulate ROS production and inhibit MMP collapse and B cell death.
These results provide additional evidence that BAFF or Syk may be valuable targets for B cell-mediated autoimmune disease therapies, such as the chemical KR3342636. REFERENCES * Mackay, F.
& Browning, J. L. BAFF: a fundamental survival factor for B cells. _Nat. Rev. Immunol._ 2, 465–475. https://doi.org/10.1038/nri844 (2002). Article CAS PubMed Google Scholar * Moon,
E. Y. & Ryu, S. K. TACI: Fc scavenging B cell activating factor (BAFF) alleviates ovalbumin-induced bronchial asthma in mice. _Exp. Mol. Med._ 39, 343–352.
https://doi.org/10.1038/emm.2007.38 (2007). Article CAS PubMed Google Scholar * Reyes, L. I. _et al._ Dexamethasone inhibits BAFF expression in fibroblast-like synoviocytes from patients
with rheumatoid arthritis. _Cytokine_ 42, 170–178. https://doi.org/10.1016/j.cyto.2007.12.005 (2008). Article CAS PubMed Google Scholar * Mackay, F. & Schneider, P. Cracking the
BAFF code. _Nat. Rev. Immunol._ 9, 491–502. https://doi.org/10.1038/nri2572 (2009). Article CAS PubMed Google Scholar * Schneider, P. _et al._ BAFF, a novel ligand of the tumor necrosis
factor family, stimulates B cell growth. _J. Exp. Med._ 189, 1747–1756 (1999). Article CAS Google Scholar * Moore, P. A. _et al._ BLyS: Member of the tumor necrosis factor family and B
lymphocyte stimulator. _Science_ 285, 260–263. https://doi.org/10.1126/science.285.5425.260 (1999). Article CAS PubMed Google Scholar * Gross, J. A. _et al._ TACI-Ig neutralizes
molecules critical for B cell development and autoimmune disease. Impaired B cell maturation in mice lacking BLyS. _Immunity_ 15, 289–302 (2001). Article CAS Google Scholar * Swee, L. K.,
Tardivel, A., Schneider, P. & Rolink, A. Rescue of the mature B cell compartment in BAFF-deficient mice by treatment with recombinant Fc-BAFF. _Immunol. Lett._ 131, 40–48.
https://doi.org/10.1016/j.imlet.2010.03.007 (2010). Article CAS PubMed Google Scholar * Khare, S. D. _et al._ Severe B cell hyperplasia and autoimmune disease in TALL-1 transgenic mice.
_Proc. Natl. Acad. Sci. USA_ 97, 3370–3375. https://doi.org/10.1073/pnas.050580697 (2000). Article ADS CAS PubMed Google Scholar * Affara, N. I. _et al._ B cells regulate macrophage
phenotype and response to chemotherapy in squamous carcinomas. _Cancer Cell_ 25, 809–821. https://doi.org/10.1016/j.ccr.2014.04.026 (2014). Article CAS PubMed PubMed Central Google
Scholar * Mittal, M., Siddiqui, M. R., Tran, K., Reddy, S. P. & Malik, A. B. Reactive oxygen species in inflammation and tissue injury. _Antioxid. Redox Signal._ 20, 1126–1167.
https://doi.org/10.1089/ars.2012.5149 (2014). Article CAS PubMed PubMed Central Google Scholar * Kawagishi, H. & Finkel, T. Unraveling the truth about antioxidants: ROS and disease:
finding the right balance. _Nat. Med._ 20, 711–713. https://doi.org/10.1038/nm.3625 (2014). Article CAS PubMed Google Scholar * Tohyama, Y., Takano, T. & Yamamura, H. B cell
responses to oxidative stress. _Curr. Pharm. Des._ 10, 835–839 (2004). Article CAS Google Scholar * Halliwell, B. & Gutteridge, J. M. Role of free radicals and catalytic metal ions in
human disease: an overview. _Methods Enzymol._ 186, 1–85 (1990). Article CAS Google Scholar * Gottlieb, E., Armour, S. M., Harris, M. H. & Thompson, C. B. Mitochondrial membrane
potential regulates matrix configuration and cytochrome c release during apoptosis. _Cell Death Differ._ 10, 709–717. https://doi.org/10.1038/sj.cdd.4401231 (2003). Article CAS PubMed
Google Scholar * Marchi, S. _et al._ Mitochondria-ros crosstalk in the control of cell death and aging. _J. Signal Transduct._ 2012, 329635. https://doi.org/10.1155/2012/329635 (2012).
Article CAS PubMed Google Scholar * Jonckheere, A. I., Smeitink, J. A. & Rodenburg, R. J. Mitochondrial ATP synthase: architecture, function and pathology. _J. Inherit. Metab. Dis._
35, 211–225. https://doi.org/10.1007/s10545-011-9382-9 (2012). Article CAS PubMed Google Scholar * Salvioli, S., Ardizzoni, A., Franceschi, C. & Cossarizza, A. JC-1, but not DiOC6(3)
or rhodamine 123, is a reliable fluorescent probe to assess delta psi changes in intact cells: implications for studies on mitochondrial functionality during apoptosis. _FEBS Lett._ 411,
77–82 (1997). Article CAS Google Scholar * Patke, A., Mecklenbrauker, I., Erdjument-Bromage, H., Tempst, P. & Tarakhovsky, A. BAFF controls B cell metabolic fitness through a PKC
beta- and Akt-dependent mechanism. _J. Exp. Med._ 203, 2551–2562. https://doi.org/10.1084/jem.20060990 (2006). Article CAS PubMed PubMed Central Google Scholar * Moon, E. Y., Ryu, S. K.
& Oh, S. Y. Reactive oxygen species (ROS) augment B-cell activating factor (BAFF) expression. _Febs. J._ 273, 149–149 (2006). Google Scholar * Turner, M. _et al._ Perinatal lethality
and blocked B-cell development in mice lacking the tyrosine kinase Syk. _Nature_ 378, 298–302. https://doi.org/10.1038/378298a0 (1995). Article ADS CAS PubMed Google Scholar * Cheng, A.
M. _et al._ Syk tyrosine kinase required for mouse viability and B-cell development. _Nature_ 378, 303–306. https://doi.org/10.1038/378303a0 (1995). Article ADS CAS PubMed Google
Scholar * Lupher, M. L. Jr. _et al._ Cbl-mediated negative regulation of the Syk tyrosine kinase. A critical role for Cbl phosphotyrosine-binding domain binding to Syk phosphotyrosine 323.
_J. Biol. Chem._ 273, 35273–35281 (1998). Article CAS Google Scholar * Simon, M., Vanes, L., Geahlen, R. L. & Tybulewicz, V. L. Distinct roles for the linker region tyrosines of Syk
in FcepsilonRI signaling in primary mast cells. _J. Biol. Chem._ 280, 4510–4517. https://doi.org/10.1074/jbc.M410326200 (2005). Article CAS PubMed Google Scholar * Yankee, T. M.,
Keshvara, L. M., Sawasdikosol, S., Harrison, M. L. & Geahlen, R. L. Inhibition of signaling through the B cell antigen receptor by the protooncogene product, c-Cbl, requires Syk tyrosine
317 and the c-Cbl phosphotyrosine-binding domain. _J. Immunol._ 163, 5827–5835 (1999). CAS PubMed Google Scholar * Mocsai, A., Ruland, J. & Tybulewicz, V. L. The SYK tyrosine kinase:
a crucial player in diverse biological functions. _Nat. Rev. Immunol._ 10, 387–402. https://doi.org/10.1038/nri2765 (2010). Article CAS PubMed PubMed Central Google Scholar * Zhang,
J., Billingsley, M. L., Kincaid, R. L. & Siraganian, R. P. Phosphorylation of Syk activation loop tyrosines is essential for Syk function. An in vivo study using a specific anti-Syk
activation loop phosphotyrosine antibody. _J. Biol. Chem._ 275, 35442–35447. https://doi.org/10.1074/jbc.M004549200 (2000). Article CAS PubMed Google Scholar * Qin, S. _et al._
Cooperation of tyrosine kinases p72syk and p53/56lyn regulates calcium mobilization in chicken B cell oxidant stress signaling. _Eur. J. Biochem._ 236, 443–449 (1996). Article CAS Google
Scholar * Qin, S., Minami, Y., Hibi, M., Kurosaki, T. & Yamamura, H. Syk-dependent and -independent signaling cascades in B cells elicited by osmotic and oxidative stress. _J. Biol.
Chem._ 272, 2098–2103 (1997). Article CAS Google Scholar * Patterson, H. C. _et al._ A respiratory chain controlled signal transduction cascade in the mitochondrial intermembrane space
mediates hydrogen peroxide signaling. _Proc. Natl. Acad. Sci. USA_ 112, E5679–E5688. https://doi.org/10.1073/pnas.1517932112 (2015). Article CAS PubMed Google Scholar * Schweighoffer, E.
_et al._ The BAFF receptor transduces survival signals by co-opting the B cell receptor signaling pathway. _Immunity_ 38, 475–488. https://doi.org/10.1016/j.immuni.2012.11.015 (2013).
Article CAS PubMed PubMed Central Google Scholar * Wang, W. H., Childress, M. O. & Geahlen, R. L. Syk interacts with and phosphorylates nucleolin to stabilize Bcl-x(L) mRNA and
promote cell survival. _Mol. Cell Biol._ 34, 3788–3799. https://doi.org/10.1128/MCB.00937-14 (2014). Article CAS PubMed PubMed Central Google Scholar * Hobeika, E. _et al._ CD19 and
BAFF-R can signal to promote B-cell survival in the absence of Syk. _EMBO J._ 34, 925–939. https://doi.org/10.15252/embj.201489732 (2015). Article CAS PubMed PubMed Central Google
Scholar * Paiva, C. _et al._ SYK inhibition thwarts the BAFF—B-cell receptor crosstalk and thereby antagonizes Mcl-1 in chronic lymphocytic leukemia. _Haematologica_ 102, 1890–1900.
https://doi.org/10.3324/haematol.2017.170571 (2017). Article CAS PubMed PubMed Central Google Scholar * Jang, J. W., Lee, J. W., Yoon, Y. D., Kang, J. S. & Moon, E. Y. Bisphenol A
and its substitutes regulate human B cell survival via Nrf2 expression. _Environ. Pollut._ 259, 113907. https://doi.org/10.1016/j.envpol.2019.113907 (2020). Article CAS PubMed Google
Scholar * Lee, G. H. _et al._ KR33426, [2-(2,5-dichlorophenyl)-5-methyloxazol-4yl]carbonylguanidine, is a novel compound to be effective on mouse systemic lupus erythematosus. _Eur. J.
Pharmacol._ 668, 459–466. https://doi.org/10.1016/j.ejphar.2011.07.026 (2011). Article CAS PubMed Google Scholar * Lee, J. W., Park, S., Kim, S. Y., Um, S. H. & Moon, E. Y. Curcumin
hampers the antitumor effect of vinblastine via the inhibition of microtubule dynamics and mitochondrial membrane potential in HeLa cervical cancer cells. _Phytomedicine_ 23, 705–713.
https://doi.org/10.1016/j.phymed.2016.03.011 (2016). Article CAS PubMed Google Scholar * Charles, I. _et al._ Serum deprivation induces apoptotic cell death of transformed rat retinal
ganglion cells via mitochondrial signaling pathways. _Invest. Ophthalmol. Vis. Sci._ 46, 1330–1338. https://doi.org/10.1167/iovs.04-0363 (2005). Article PubMed Google Scholar * Lee, S. B.
_et al._ Serum deprivation-induced reactive oxygen species production is mediated by Romo1. _Apoptosis Int. J. Program. Cell Death_ 15, 204–218. https://doi.org/10.1007/s10495-009-0411-1
(2010). Article CAS Google Scholar * Tait, S. W. & Green, D. R. Mitochondria and cell death: outer membrane permeabilization and beyond. _Nat. Rev. Mol. Cell Biol._ 11, 621–632.
https://doi.org/10.1038/nrm2952 (2010). Article CAS PubMed Google Scholar * Dawson, T. J., Webster, K. N., Lee, E. & Buttemer, W. A. High muscle mitochondrial volume and aerobic
capacity in a small marsupial (_Sminthopsis crassicaudata_) reveals flexible links between energy-use levels in mammals. _J. Exp. Biol._ 216, 1330–1337. https://doi.org/10.1242/jeb.079087
(2013). Article CAS PubMed Google Scholar * Ackermann, J. A. _et al._ Syk tyrosine kinase is critical for B cell antibody responses and memory B cell survival. _J. Immunol._ 194,
4650–4656. https://doi.org/10.4049/jimmunol.1500461 (2015). Article CAS PubMed PubMed Central Google Scholar * Hampe, C. S. B. Cell in autoimmune diseases. _Scientifica_
https://doi.org/10.6064/2012/215308 (2012). Article PubMed PubMed Central Google Scholar * Suurmond, J. & Diamond, B. Autoantibodies in systemic autoimmune diseases: specificity and
pathogenicity. _J. Clin. Invest._ 125, 2194–2202. https://doi.org/10.1172/JCI78084 (2015). Article PubMed PubMed Central Google Scholar * Eggert, M., Zettl, U. K. & Neeck, G.
Autoantibodies in autoimmune diseases. _Curr. Pharm. Des._ 16, 1634–1643 (2010). Article CAS Google Scholar * Pogue, S. L., Kurosaki, T., Bolen, J. & Herbst, R. B cell antigen
receptor-induced activation of Akt promotes B cell survival and is dependent on Syk kinase. _J. Immunol._ 165, 1300–1306 (2000). Article CAS Google Scholar * Lee, C. K., Yang, Y., Chen,
C. & Liu, J. Syk-mediated tyrosine phosphorylation of mule promotes TNF-induced JNK activation and cell death. _Oncogene_ 35, 1988–1995. https://doi.org/10.1038/onc.2015.275 (2016).
Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS This work was supported by the R&D program for Society of the National Research Foundation (NRF) funded by the
Ministry of Science, ICT & Future Planning (Grant from Mid-career Researcher Program: #2016R1A2B4007446 and # 2018R1A2A3075602), Republic of Korea. AUTHOR INFORMATION AUTHORS AND
AFFILIATIONS * Department of Integrative Bioscience and Biotechnology, Sejong University, 209 Neungdong-ro Kwangjin-gu, Seoul, 05006, Republic of Korea Sojin Park, Ju-Won Jang & Eun-Yi
Moon Authors * Sojin Park View author publications You can also search for this author inPubMed Google Scholar * Ju-Won Jang View author publications You can also search for this author
inPubMed Google Scholar * Eun-Yi Moon View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS S.J.P. settled down and conducted the experiments,
and wrote primary manuscript. J.-W.J. conducted the experiments and revised manuscript. E.-Y.M. planed main idea of the study, analyzed the results, corrected the manuscript, and supported
S.J.P. and J.-W.J. to provide reagents, materials and analysis tools. All authors reviewed the manuscript. CORRESPONDING AUTHOR Correspondence to Eun-Yi Moon. ETHICS DECLARATIONS COMPETING
INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and
institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the
source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative
Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by
statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Park, S., Jang, JW. & Moon, EY. BAFF attenuates oxidative stress-induced cell
death by the regulation of mitochondria membrane potential via Syk activation in WiL2-NS B lymphoblasts. _Sci Rep_ 10, 11784 (2020). https://doi.org/10.1038/s41598-020-68628-5 Download
citation * Received: 12 February 2019 * Accepted: 16 March 2020 * Published: 16 July 2020 * DOI: https://doi.org/10.1038/s41598-020-68628-5 SHARE THIS ARTICLE Anyone you share the following
link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature
SharedIt content-sharing initiative