Play all audios:
ABSTRACT Serious human health impacts have been observed worldwide due to several life-threatening diseases such as cancer, candidiasis, hepatic coma, and gastritis etc. Exploration of
nature for the treatment of such fatal diseases is an area of immense interest for the scientific community. Based on this idea, the genus _Aspergillus_ was selected to discover its hidden
therapeutic potential. The genus _Aspergillus_ is known to possess several biologically active compounds. The current research aimed to assess the biological and pharmacological potency of
the extracts of less-studied _Aspergillus ficuum_ (FCBP-DNA-1266) (_A. ficuum_) employing experimental and bioinformatics approaches. The disc diffusion method was used for the antifungal
investigation, and the MTT assay was performed to assess the anticancer effects. Mice were employed as an in vivo_ model_ to evaluate the antispasmodic effects. A standard spectrophotometric
technique was applied to gauge the urease inhibitory activity. The antifungal studies indicate that both n-hexane and ethyl acetate extracts were significantly active against _Candida
albicans_ (_C. albicans_) with their zone of inhibitions (ZOI) values reported as 19 ± 1.06 mm and 25 ± 0.55 mm, respectively at a dose of 30 µg.mL−1. In vitro cytotoxicity assay against
HeLa, fibroblast 3T3, prostate PC3, and breast MCF-7 cancer cell lines was performed. The ethyl acetate extract of _A. ficuum_ was found to be significantly active against MCF-7 with its
IC50 value of 43.88 µg.mL−1. However, no substantial effects on the percent cell death of HeLa cancer cell lines were observed. In addition, the _A. ficuum_ extracts also inhibited the
urease enzyme compared to standard thiourea. The antispasmodic activity of _A. ficuum_ extract was assessed by an in vivo model and the results demonstrated promising activity at 150
mg.kg−1. Molecular docking results also supported the antifungal, anticancer, and antiurease potency of _A. ficuum_ extract. Overall, the results display promising aspects of _A. ficuum_
extract as a future pharmacological source. SIMILAR CONTENT BEING VIEWED BY OTHERS SYNTHESIS, CHARACTERIZATION AND BIO-EVALUATION OF NOVEL SERIES OF PYRAZOLINE DERIVATIVES AS POTENTIAL
ANTIFUNGAL AGENTS Article Open access 28 April 2025 EXPLORING THE ANTIFUNGAL, ANTIBIOFILM AND ANTIENZYMATIC POTENTIAL OF ROTTLERIN IN AN IN VITRO AND IN VIVO APPROACH Article Open access 15
May 2024 NON-ANTIFUNGAL DRUGS INHIBIT GROWTH, MORPHOGENESIS AND BIOFILM FORMATION IN _CANDIDA ALBICANS_ Article 19 January 2021 INTRODUCTION New drugs are needed due to certain factors such
as globalization, the aging population, and microbial resistance to existing medicines. Only a small number of biologically active compounds are currently used in prescription drugs1.
Scientists are currently investigating emerging diseases to gain a better understanding of them and find their potential cures using various natural and chemical formulations. However, there
are still many areas that remain unexplored due to insufficient knowledge and techniques. To find new bioactive compounds that can be used for medicinal, agricultural, and industrial
purposes, researchers are focusing on isolating such compounds from new species of fungi2. In recent times, researchers have isolated several metabolites from fungi, including
anthraquinones, alkaloids, terpenes, steroids, flavonoids, and cyclic peptides. Some secondary metabolites produced by various fungi have been found to exhibit potent biological activities
such as antifungal, antitumor, anti-inflammatory antibacterial, antiparasitic, and antiviral properties. The chemical diversity of fungal secondary metabolites provides a significant
advantage for the development of new drugs3,4. Anofinic acid, a potent active metabolite produced by _Aspergillus tubingensis_, has shown strong antimicrobial activity against a variety of
hazardous bacteria in addition to its potent activity against various carcinoma cells, including Hep-G2, MCF-7, and HCT-1162. Sorbicillinoids possess the capability to be used as
pharmaceutical and agrochemical agents due to their antiviral, antimicrobial, and anticancer effects besides their use as food colorants, and pigments. Sorbicillin and sorbicatechol D have
been reported to exhibit antiproliferative effects against HT-29 tumor cells dose-dependently5. The metabolites gliotoxin, deoxytryptoquivaline, and patulin isolated from various
_Aspergillus_ species are known to induce apoptosis in cells6,7,8. A study performed on the secondary metabolites (terretonin N and butyrolactone I) of _A. terreus_ confirmed their promising
anticancer potential. Cellular apoptosis was induced at higher rates in cancer cells lacking a necrotic apoptotic pathway9. Similarly, recent studies on extracts of various fungi,
particularly _Aspergillus,_ have revealed their potent anticancer, antinociceptive, and anti-inflammatory activities10,11. In addition, the latest studies have also confirmed the strong
enzyme-inhibiting effects of numerous species of fungi9. _Aspergillus ficuum_ is a filamentous fungus that belongs to the _Niger_ clade in the _Aspergillus_ section _Nigri_. Its
morphological characteristics are the formation of spores and the display of colony characteristics. The taxonomy of the _Niger_ clade depends on the generation of various secondary
metabolites belonging to the five classes such as pyranone, alkaloid, cyclopentapeptide, polyketide, and sterol. These metabolites are known as naphthopyrones, malformins, bicoumarins
(kotanins), fumonisin B2 and B4, citric acid, diketopiperazine, asperazine, and other related compounds12. Extensive biological aspects have been reported for the _Niger_ clade13. In our
previous study, we investigated the metabolic profile of _A. ficuum_ extracts for the first time, which led to the discovery of various bioactive compounds, including hydroxyvittatine,
aurasperone D, choline sulfate, noruron, cetrimonium, heneicosane, kurilensoside, eicosane, and nonadecane. Furthermore, a pharmacological analysis of _A. ficuum_ extracts was also carried
out via in vitro and in vivo models14. In continuation of our previous study, extracts of the lesser-explored member of the Niger clade, _A. ficuum_ were further investigated for their
antifungal and anticancer activities via an in vitro model. Whereas their anti-inflammatory and antispasmodic effects were studied through an in vivo model. In addition, computational
analyses were performed to support the in vitro and in vivo results. These investigations of _A. ficuum_ extracts will offer new understandings into the area of the fungal diaspora.
MATERIALS AND METHODS CHEMICALS AND ANIMALS All standard chemicals and solvents were obtained from Sigma Aldrich. The (BALB/c) mice weighing 25–35 g were purchased from the National
Institute of Health (NIH), Islamabad, Pakistan. They were acclimated at 22 °C on a light/dark cycle (12 h/12 h) with adequate provision of water and food throughout the study. All methods
were performed following the relevant guidelines and regulations15. The in vivo studies were conducted under the number 7196/LM/UoA Ethical Committee FAHV&S, the University of
Agriculture Peshawar, Pakistan14. FUNGAL STRAIN The fungal strain (_A. ficuum_, FCBP-DNA-1266) was obtained from the fungal bank of the University of Punjab, Pakistan. CULTURE CULTIVATION
The fungal culture was cultured at 28 °C for 21 days in the static state. _A. ficuum_ spores (105 conidia/mL) were transferred to the multiple Erlenmeyer flask (500 mL) containing about 250
mL of Potato Dextrose Broth (PDB). To inhibit bacterial growth in the medium, PDB was added with 25 mg.L−1 of streptomycin sulfate16. EXTRACTION AND FRACTIONATION After 21 days, the mycelia
formed in each flask were removed and processed for drying. The mycelia were washed several times with deionized water to remove any media ingredients. The constant dry weight of mycelia was
obtained after drying at 60 °C in an oven. The dried mycelia were crushed to powder using a mortar and pestle. The powdered mycelia weighing 60 g was extracted in triplicate using three
hundred milliliters of ethyl acetate (EtOAc) (3 × 300 mL). Subsequently, the obtained 4 g of ethyl acetate extract was fractionated by n-hexane (3 × 300 mL). Both portions were condensed via
a rotary evaporator (Buchi, Germany, Model R-300). The dried-up portions, ethyl acetate (2.3 g) and n-hexane (1.7 g) were kept at 4 °C in the refrigerator for further studies16,17.
ANTIFUNGAL ASSAY The microorganisms such as _Aspergillus niger_ (_A. niger_), _Fusarium oxysporum_ (_F. oxysporum_), _Trichoderma harzianum_ (_T. harzianum_)_, Candida albicans_ (_C.
albicans_)_,_ and _Aspergillus flavus_ (_A. flavus_) used for the antifungal assay were acquired from the Department of Agricultural Chemistry and Biochemistry, the University of
Agriculture, Peshawar, Pakistan. A sterilized malt extract agar (MEA) medium was prepared and poured into Petri dishes. The spores from each fungal inoculum were added to these plates. The
sterilized discs were soaked in _A. ficuum_ extracts at concentrations of 10, 20, and 30 µg.mL−1 and were then positioned on the plates. Subsequently, the plates were cultured at 28 °C and
then analyzed for antifungal assay after 72 h. Mancozeb was used as a drug control while DMSO was utilized as a negative control. The antifungal analysis was performed by measuring the zone
of inhibition in millimeters17. ANTISPASMODIC ACTIVITY Standard protocol was followed with minor modifications18,19. Mice weighing 20–30 g were used for the antispasmodic activity. Before
the experiments, five mouse groups were created and kept on fast. Ethyl acetate extract of _A. ficuum_ was introduced in triplicate at a dose of 10–20 mg.Kg−1. Two groups were orally
administered castor oil (0.5 mL) and normal saline water (negative control) at a dose of 10 mL.kg−1. The residual three groups were orally administered _A. ficuum_ extract at a dose of 10–20
mg.kg−1. Each group of mice was orally administered with charcoal (1 mL) after 1 h of induction of castor oil, and extract. The passage of charcoal from the pylorus to the caecum was noted
by sacrificing mice through cervical dislocation after 50 min of charcoal induction. The percent inhibition caused by the _A. ficuum_ extract was determined as under: Percent inhibition =
[Distance covered by charcoal/Total length of intestine] × 100. MTT ASSAY The anticancer studies were carried out at Hussain Ejaz Institute, Department of Chemistry, University of Karachi. A
standard colorimetric MTT (3-[4, 5-dimethylthiazole-2-yl]-2, 5-diphenyl-tetrazolium bromide) analysis was performed to determine the cytotoxic potential of crude extract of _A. ficuum_ by
using 96-well flat-bottomed microplates20. In this study, HeLa cells (Cervical Cancer), 3T3 (mouse fibroblast), PC3 cells (Prostrate Cancer), and MCF-7 (Breast Cancer) cells were examined.
These different cancer cells were cultivated in Minimum Essential Medium Eagle (MEME), added with fetal bovine serum (5%) (FBS), penicillin (100 IU.mL−1), and streptomycin (100 µg.mL−1) in
flasks (75 cm2). The flasks were cultured at 37 °C in a CO2 (5%) incubator. The substantially growing cells were counted with a hemocytometer followed by dilution with a specific medium. A
volume of 100 µL of each cell culture (HeLa, 3T3, MCF-7, and PC3) having 1 × 104 cells/well was seeded into 96-well plates at a concentration of 100 µL/well. The cell cultures were incubated
overnight and the medium was separated from them. A newly harvested medium (200 µL) was administered to the well of each cell culture along with different concentration doses of extract of
_A. ficuum_ (15–60 µg.mL−1). After incubation for 48 h, each well was added MTT solution (200 µL) having a concentration of 5 mg.mL−1. The cell cultures were further nurtured for 4 h. Later,
DMSO (100 µL) was introduced to each well. The synthesis of formazan by the decrease of MTT was determined using a microplate reader (Spectra Max Plus, Molecular Devices, CA, USA) through
absorbance maximum at a wavelength of 570 nm. Doxorubicin and DMSO were used as a positive and negative control, respectively. The 50% growth inhibition (IC50) was measured for cytotoxicity
by using the straight-line equation y = mx + c. By using Soft-Max Pro software (Molecular Device, USA), the following formula was applied for the measurement of percent inhibition: %
inhibition = 100 − (mean of O.D of _A ficuum_ extract − mean of O.D of negative control)/(mean of O.D of positive control – mean of O.D of negative control) *100). UREASE INHIBITION ASSAY
The urease inhibitory effect of the crude extracts of _A. ficuum_ was evaluated by following the standard protocol with little modifications21. A solution of 20 µL of Jack bean urease (2.5
units/mL) and 50 µL of urea (100 mM) was prepared. The solution was dissolved in phosphate buffer, pH 8.2 [K2HPO4·3H2O (0.01 M), EDTA (1.0 m), and LiCl2 (0.01 M)] and was mixed with 30 µL of
250 µg.mL−1 concentrations of EtOAc and n-hexane extracts. The mixture was subjected to incubation at 37 °C in a 96-well plate for 10 min. The reaction mixture was incubated at 30 °C in a
96-well plate for 15 min. The indophenol method was used to determine the amount of ammonia production to measure the suppression of urease activity. The phenol reagent [phenol (1%), sodium
nitroprusside (0.005%), 40 µL] and the alkaline reagent [60 µL, NaOH (0.5%), NaOCl (0.1%)] were added to each well. Thiourea (100 µg.mL−1) was taken as a standard and data were obtained in
triplicate by measuring the absorption maximum at 630 nm. Percent inhibition was determined via the equation: Percent inhibition = 100 − (OD _A. ficuum extracts_/OD control) × 100. IN SILICO
ANALYSIS Mycocompounds (Fig. S1) tentatively reported for _A. ficuum_ extracts14 were chosen for docking assay against heat shock protein 90 (hsp90) (PDB ID: 6CJI) of _C. albicans_, jack
bean urease (PDB ID: 3LA4) and human HER2 protein (PDB ID: 3PP0) at a resolution of 1.64, 2.05 and 2.25 Å, respectively17,22,23. High-resolution X-ray crystal structures of the chosen
proteins were obtained from the Protein Data Bank (http://www.rcsb.org/pdb). By using a preparation program integrated into the Molecular Operating Environment (MOE) software, protein
molecules were optimized for docking study by eliminating water molecules, introducing missing hydrogen atoms, allocating the correct hybridization state to each atom in each residue, and
fixing charges. Active sites of the designated proteins were located using the active site finder tool embedded in MOE software. Using the docking tool of the MOE software, mycocompounds
were docked to the active binding sites of the chosen pathogenic proteins. For each molecule, 30 conformations were produced using specific torsion angles for all the rotatable bonds. The
MOE-implanted London dock scoring function was used to determine the binding energy for each protein–ligand complex system24. STATISTICAL ANALYSIS Statistical analysis of all data obtained
was performed using GraphPad Prism version 8.0. Results were evaluated using a mean n = 3 with standard deviation. Antifungal analyses were evaluated through one-way ANOVA while means were
separated by applying LSD at _P_ ≤ 0.05. Similarly, the results of the in vivo pharmacological investigation were evaluated through one-way ANOVA, followed by post hoc analysis (Dunnett’s
test). ETHICS APPROVAL All experimental protocols were approved by the ethical committee FAHV&S, the University of Agriculture Peshawar, Pakistan under the number 7196/LM/UoA. All
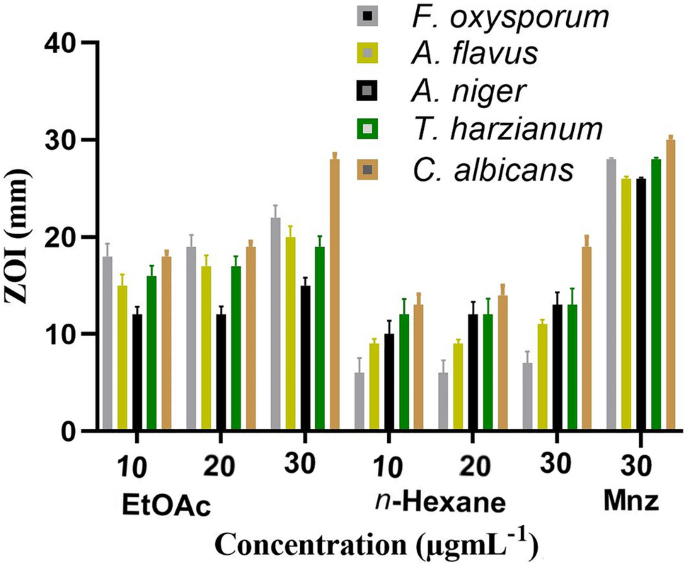
methods are reported following ARRIVE guidelines (https://arriveguidelines.org). RESULTS AND DISCUSSIONS ANTIFUNGAL ACTIVITY The antifungal analysis of ethyl acetate and n-hexane extracts of
_A. ficuum_ was determined versus five fungal species (Table 1). _C. albicans_ is a mutual fungus that causes death by colonizing the human skin and gastrointestinal tract. It is the
causative agent of multidrug resistance in individuals with compromised immune systems due to multiple diseases. Several studies have been conducted on its control using natural
products25,26. In this study, the antifungal investigations reveal that ethyl acetate extract was significantly active (_P_ ≤ 0.05) against _C. albicans_ with a ZOI of 25 ± 0.55, at a dose
of 30 µg.mL−1. It was also recorded that ethyl acetate extract was substantially active (_P_ ≤ 0.05) against all fungal species at a dose concentration of 30 µg.mL−1. Likewise, potent
antifungal activity (_P_ ≤ 0.01–0.05) was also exhibited by n-hexane extract against other fungal species (Fig. 1). The n-hexane extract was also profoundly active (_P_ ≤ 0.05) against the
tested fungal strains at high concentrations. The fungal species are known to be great synthesizers of antifungal compounds. Several classes of compounds produced by the genus _Aspergillus_,
such as statins and polyketides, are considered effective antifungal compounds27. A study conducted on the extract isolated from the endophytic fungus _Colletotrichum gloeosporioides_
displayed potent antifungal activity against _C. albicans_28. In addition, a similar study was conducted on various fungal strains of different genera such as _Acremonium_, _Fusarium_,
_Aspergillus,_ and _Penicillium_ which showed potent antifungal activities29. In another study, the n-hexane extract was found to be effective against _A. flavus_ with a ZOI of 21.63 mm and
less significant against _F. oxysporum_ with a ZOI of 15.31 mm. The genus _Aspergillus_ is considered a good producer of antimicrobial agents without any significant toxicity30,31.
ANTISPASMODIC ACTIVITY Inflammatory bowel disorders and gastritis are very common in humans and are induced by histamine and acetylcholine. The gastrointestinal tract becomes irritated and
inflamed by smooth muscle contractions, causing uneasiness and discomfort. Such conditions are usually treated with antispasmodics which also show numerous side effects18. A charcoal
motility assay was performed to assess the inhibition of ethyl acetate extract of _A. ficuum_ (Table 2). The three different dosages of extracts from _A. ficuum_ were able to show an
antispasmodic effect. Smooth muscle contractions were reduced in a dose-dependent manner by ethyl acetate extract compared to mice treated with castor oil. It was observed that as
concentrations of _A. ficuum_ extract increased, there was a gradual increase in spasmodic activity, which is not a good indication. However, _A. ficuum_ extracts possessed antispasmodic
activity compared to the percent mobility of charcoal in castor oil, providing evidence that _A. ficuum_ extracts can relieve spasms (Fig. 2). According to the literature, an antispasmodic
effect against three different spasmogens was shown in a study of the n-hexane extract from endophytic fungi32. The literature review showed that bio-transformed diterpenes from _A. niger_
displayed significant antispasmodic activity33. Although this study was conducted using a few concentrations of the extract, further investigation of the mycochemicals of this fungus will
lead to the development of the most standardized and effective drug. ANTICANCER ACTIVITY The anticancer activity of ethyl acetate extracts of _A. ficuum_ was measured versus HeLa, 3T3, PC3,
and breast cancer cells (MCF-7) (Table 3). In the case of MCF-7, the IC50 value was noted to be 43.88 µg.mL−1, significantly potent compared to other cancer cell lines, HeLa, 3T3 & PC3
with their IC50 noted to be 250.1, 75.54, and 70.07 µg.mL−1, respectively. From the analysis, it was visible that percent cell viability decreases with the increase in the concentration of
the extract. The linear pattern of cell viability against the dose concentrations of the extracts was evident (Fig. 3). At a high dose of 60 µg.mL−1, no significant difference was observed
for the positive control, doxorubicin. A similar pattern was observed for all doses of _A. ficuum_ extract against the 3T3 cancer cell line. Moreover, no significant effect of _A. ficuum_
extract was found against the HeLa cell line. It was also evident that high doses of _A. ficuum_ extract caused significant cell death (Fig. 4). Previously, a study showed that more than 100
anticancer drugs associated to 19 diverse chemical groups were obtained from fungi34. Artika et al_._ studied the anticancer activity of endophytic fungi. Their data showed that only one
isolated chemical at a concentration of 400 µg.mL−1 exhibited potency comparable to the standard drug35. Felczykowska and coworkers found that fungi possess antiproliferative materials. They
studied that the fungus, _Protoparmeliopsis muralis_ isolated from lichen exhibited potent activity versus MCF-7 cancer cells36. Tincho et al_._ studied the cytotoxic potential of numerous
fungi isolated from _Terminalia cattapa_. They discovered that an extract of _F. oxysporum_ exhibited a potent 50 percent inhibitory potential versus cancer cell lines37. Thomas et
al.examined twenty-one fungal extracts versus MCF-7 cancer cells. Their results revealed that sample F-21 possessed strong anticancer activity with its IC50 value of 44.75 µg.mL−138.
Nevertheless, several species of _Aspergillus_ are highlighted as potent producers of anticancer metabolites39. The biosynthesis of secondary metabolites in fungi is highly reliant on the
substrate and other ecological conditions40. Since this study was performed on a PDB medium, mycelial growth on other substrates and optimized conditions could contribute to the potent
anticancer activities of fungal extracts. UREASE INHIBITION ASSAY The conversion of urea into harmful products by the urease enzyme has serious effects on plants, animals, and humans. Its
action causes urinary stones, gastritis, gastric cancer, hepatic coma, and other serious diseases in living organisms. The ureolytic activity of urease was suppressed by the natural products
obtained from various sources41. The Urease inhibition assay of different extracts of _A. ficuum_ was assessed in this study. A significant percent urease inhibitory potential was exhibited
by ethyl acetate extract of _A. ficuum_. The ethyl acetate and n-hexane extracts of _A. ficuum_ suppressed urease by 71.58 and 53.22%, respectively. The result agreed well with standard
thiourea (Fig. 5). The difference between the ethyl acetate extract and the standard thiourea was not significant, which is a good sign as the commercially available ureases are toxic and
less stable. The study regarding the suppression of the urease enzyme by fungi is very limited. Rauf et al.determined that the ethyl acetate fraction of _Screlotium rolfsii_ and _A. flavus_
was significantly inactive to urease enzymes with less than 50% inhibition42. A literature review suggested that fungal metabolites can inhibit the urease enzyme at different dose levels43.
A research group isolated two compounds from _Paecilomyces formosus_ which exhibited significant urease inhibitory activity with their potencies of 75.8 and 190.5 µg.mL−144. Another group of
researchers found the urease inhibitory activity of a metabolite isolated from fungi, Bipolaris sorokiniana LK12. In a dose-dependent study, the metabolites showed an IC50 value of 81.62
µg.mL−1 against the urease enzyme45. DOCKING ANALYSIS Ethyl acetate extract of _A. ficuum_ was found significantly active against all tested fungi in general and _C. albicans_ in particular.
Therefore, the mycocompounds were cautiously identified from ethyl acetate extract14 [Fig. S1] were docked versus the Hsp90 protein of _C. albicans_ to support the antifungal activity of
_A. ficuum_ extract. Hsp90 plays a vital role in pathogenesis by performing protein biogenesis as well as interacting with various cellular proteins46 and is referred to as a molecular
chaperone; its inhibition will halt fungal infection. Docking results reveal that all selected ligands (L1-L9) can inhibit the Hsp90 protein by forming a stable protein–ligand complex. Among
all docked ligands, L3 was found to be more efficient since it developed two physical interactions with the catalytic residues of Hsp90 resulting in the highest binding energy of − 7.9738
kcal.mol−1 (Table 4). One conventional H-bond was generated between residue Lys47 and the polar oxygen atom of L3; the second Pi-cation interaction was formed between the benzene ring of L3
and residue Lys47 (Fig. 6). L1 formed two physical interactions with Hsp90 causing a binding affinity of − 5.7161 kcal.mol−1 (Table 4). Catalytic residues Gly86 and Lys47 were involved in
developing two conventional H-bonds with two different hydroxyl groups of L1 (Fig. S2a,b). Ligand L2 was associated with amino acid Asn95 of Hsp90 through one H-bond; it resulted in the
binding energy of − 5.8924 kcal.mol−1 (Table 4) (Fig. S2c,d). L4 formed a stable protein–ligand complex by developing two conventional H-bond interactions; two different oxygen atoms of the
sulfonate group of L4 were found associated with the catalytic residue Lys47 (Fig. S2e,f). The binding energy was noted to be − 4.8836 kcal.mol−1 (Table 4). L5 bound to the active sites of
Hsp90 generated the second-highest binding affinity of − 7.8284 kcal.mol−1 (Table 4) among all screened ligands. Residue Lys47 developed an H-bond interaction with the carbonyl oxygen atom
of L5; the second H-bond originated between residue Lys47 and the NH group of L5 (Fig. S2g,h). L6 was linked with Hsp90 through two H-bond interactions originating binding affinity of −
6.7134 kcal.mol−1 (Table 4). Residue Asp43 formed two H-bonds with two different hydroxyl groups of L6 (Fig. S2i,j). Ligand L7 formed a stable protein–ligand complex through three H-bond
interactions. The Hydroxyl moiety of L7 participated in developing H-bond interaction with residue Glu36. Two different carbonyl groups of L7 formed two H-bond interactions with residue
Lys47 and Gly126, respectively (Fig. S2k,l). The binding energy value was calculated to be − 7.5397 kcal.mol−1 (Table 4). L8 formed two physical interactions with Hsp90 originating with the
third highest binding affinity of -7.7921 kcal.mol−1 (Table 4). One H-bond was developed between residue Asp91 and the hydroxyl moiety of L8; a second H-bond was generated between the
carbonyl oxygen atom of L8 and residue Lys47 (Fig. S2m, n). In the case of L9, two physical forces were responsible for the formation of a protein–ligand complex; the binding energy value
was noted to be − 7.0975 kcal.mol−1 (Table 4). Catalytic residue Asp91 generated two conventional H-bond interactions with L9 (Fig. S2o,p). Docking data display three major findings:
firstly, all ligands (L1-L9) can bind to catalytic residues located in the active pocket of Hsp90; secondly, L3 has the highest binding affinity with Hsp90 among all screened ligands;
thirdly, residue Lys47 is actively involved in binding to six ligands. Keeping in view the chemistry of Lys47, future drug candidates can be designed. These findings support the antifungal
activity of _A. ficuum_ extracts_._ Similarly, ligands (L1-L9) were docked against the HER2 protein to support the anticancer activity of _A. ficuum_ extracts_._ HER2 is a membrane tyrosine
kinase enzyme; it plays a major part in tumor growth and the spreading of breast cancer on activation47. Docking results indicate that ligand L6 showed the highest affinity toward the HER2
protein among all screened ligands. It is associated with HER2 protein through four H-Bond interactions resulting in the highest binding energy of − 7.8907 kcal.mol−1 (Table 5). Residue
Ser728 developed two H-Bond interactions with the same hydroxyl group of L6; each amino acid Asn850 and Asp808 formed a single H-Bond interaction with two different hydroxyl groups of L6
(Fig. 7). Ligand L1 generated three physical interactions with HER2 protein stemming binding affinity of − 5.2498 kcal.mol−1 (Table 5). Catalytic residue Asp808 participated in developing
two H-Bond interactions with the same hydrogen atom of the hydroxyl moiety of L1. Residue Ser728 formed Pi-H interaction with the benzene ring of L1 (Fig. S3a,b). L2 developed two
non-covalent interactions with HER2 protein; these interactions originated a binding affinity of − 5.1722 kcal.mol−1 (Table 5). Each of the residues (Cys805 and Ser728) formed a single
H-bond with the polar hydrogen atom of the NH moiety and the carbonyl oxygen atom of L2, respectively (Fig. S3c,d). Ligand L3 formed a stable protein–ligand complex by developing two
physical interactions with binding affinity reported to be − 6.8174 kcal.mol−1 (Table 5). Each of the amino acids (Leu726 and Cys805) generated a single H-Bond with the polar hydrogen of the
hydroxyl moiety and carbonyl oxygen atom of L3, respectively (Fig. S3e,f). L4 generated a single interaction with the HER2 protein resulting in a binding affinity of − 6.1479 kcal.mol−1
(Table 5). Residue Thr862 formed a single H-Bond interaction with the polar hydrogen of the CH2 group of L4 (Fig. S3g,h). Ligand L5 was associated with HER2 protein through two non-covalent
interactions; these forces generated the second-highest binding energy of − 7.7000 kcal.mol−1 (Table 5). Catalytic residue Ser728 formed a single H-Bond interaction with the oxygen atom of
the methoxy group of L5; residue Gly727 developed a single Pi-H interaction with the pyrrole moiety of L5 (Fig. S3i,j). Ligand L7 formed a stable protein–ligand complex by generating two
physical interactions; it resulted in a binding energy of − 6.5073 kcal.mol−1 (Table 5). Residue Ser728 originated two H-Bond interactions with the polar hydrogen atom of the hydroxyl group
and carbonyl oxygen atom of L7; Lys724 developed a single H-Bond with the hydroxyl moiety of L7 (Fig. S3k,l). L8 displayed single H-Bond interaction with HER2 protein producing the third
highest binding affinity of − 6.8651 kcal.mol−1 (Table 5). The Hydroxyl moiety of L8 was involved in the creation of the protein–ligand complex (Fig. S3m,n). Ligand L9 generated two physical
interactions with HER2 protein; these non-covalent forces resulted in a binding energy value of − 6.7778 kcal.mol−1 (Table 5). Residue Asp808 developed H-Bond and Pi-Cation interactions
with two different methyl groups of L9, respectively (Fig. S3o,p)48. In the case of HER2 protein, three new aspects emerged: firstly, all ligands (L1-L9) form a stable protein–ligand
complex; secondly, ligand L6 has a greater ability to bind to the active pocket of HER2 protein among all tested ligands; thirdly, the catalytic role of residue Ser728 is found prominently
by binding to five ligands. Structural features of L6 and residue Ser728 should be considered for drugs designed against breast cancer. Herein, docking results support the anticancer
activity of _A. ficuum_ extracts. Similarly, ligands (L1-L9) were also screened against the urease enzyme through the molecular docking technique; the results indicated that all ligands
could inhibit the urease enzyme by forming a stable protein–ligand complex. Based on binding affinity, L8 was found to have the highest binding affinity of − 7.7921 kcal.mol−1 (Table 6)
among all selected ligands. It developed four physical interactions with urease protein. Both residues (His593 and His585) generated a single Pi–Pi interaction with two different benzene
rings of L8; in addition, two residues namely Met637 and Cme592 also formed a single H-Bond interaction with the hydroxyl moiety and carbonyl oxygen atom of L8, respectively (Fig. 8). L1
originated two non-covalent interactions with the urease enzyme resulting in the binding energy of − 5.0588 kcal.mol−1 (Table 6). Residue Arg835 developed a single H-Bond interaction with
the hydroxyl moiety of L1; Arg575 formed a single H-Bond interaction with the methyl moiety of the heterocyclic ring of L1 (Fig. S4a,b). Ligand L2 generated a single non-covalent interaction
with the urease enzyme; it resulted in a binding energy value of − 5.4100 kcal.mol−1 (Table 6). Catalytic residue Val831 participated in the development of a single H-Bond interaction with
the carbonyl oxygen of L2 (Fig. S4c,d). L3 formed a stable protein–ligand complex by developing two physical interactions resulting in the binding affinity of − 6.9148 kcal.mol−1 (Table 6).
Residue Arg835 generated Pi-H and Pi-Cation interactions with the benzene ring and oxygen heterocyclic ring of L3, respectively (Fig. S4e,f). L4 generated four physical forces with the
urease enzyme; these forces resulted in a binding affinity of − 4.5903 kcal.mol−1 (Table 6). Residue Glu584 developed two H-Bond interactions with two different methyl groups of L4. Gln649
formed a single H-Bond interaction with the oxygen atom of L4 (Fig. S4g,h). Ligand L5 was involved in complex formation with urease through two physical interactions; these interactions
originated the second-highest binding affinity of − 7.6472 kcal.mol−1 (Table 6). Both residues (Glu642 and Val831) developed single H-Bond interactions with OH and NH groups of L5,
respectively (Fig. S4i,j). Ligand L6 developed a single non-covalent interaction with the active spot of the urease enzyme yielding a binding affinity of − 7.1083 kcal.mol−1 (Table 6).
Residue Arg835 displayed a single H-Bond interaction with the hydroxyl moiety of L6 (Fig. S4k,l). L7 showed three physical interactions with catalytic residues of urease enzyme generating
the third highest binding energy of − 7.5593 kcal.mol−1 (Table 6). Two residues, namely, Ser645 and Thr829 formed single H-bond interaction with two different hydroxyl groups of L7;
catalytic residue Arg835 displayed Pi-H interaction with the benzene ring of L7 (Fig. S4m,n). Ligand L9 was noticed attached to the active site of urease through two physical interactions
that resulted in a binding affinity value of − 6.7125 kcal.mol−1 (Table 6). Catalytic residue Glu642 showed two H-Bond interactions with two different methyl groups of L9 (Fig. S4o,p). Some
interesting findings were obtained from docking data: all ligands form a stable protein–ligand complex with urease enzyme; L8 shows the highest binding affinity among all screened ligands
and catalytic residue Arg835 is noticed to be more active by forming a protein–ligand complex with four ligands. CONCLUSION In this research, urease inhibition, antifungal, anticancer, and
antispasmodic activities of _A. ficuum_ extracts were explored for the first time by applying in vivo and in vitro models followed by in silico models. Like other fungi, both n-hexane and
ethyl acetate extracts of _A. ficuum_ were noticed active against five tested fungal strains. However, both extracts displayed significant activity against the _C. albicans._ Potent
antispasmodic, as well as urease inhibitory activities, were recorded for _A. ficuum_ extracts_._ Similarly, the promising anticancer effect of _A. ficuum_ against PC3, 3T3, and especially
MCF-7 cancer cell lines was documented. Furthermore, the antiurease, antifungal, and anticancer activities of _A. ficuum_ extracts were supported by molecular docking results. Among all
docked ligands, L3, L6, and L8 displayed the highest binding affinity for Hsp90, HER2, and urease proteins, respectively. Similarly residues namely, Lys47, Ser728, and Arg835 in the active
pockets of Hsp90, HER2, and urease proteins, respectively emerged as catalytically active residues generating H-Bond interactions with a greater number of ligands. Keeping in view the
structural features of ligands (L3, L6, and L8) and residues (Lys47, Ser728, and Arg835), potential drug candidates could be designed in the future for the inhibition of pathogenic proteins
such as Hsp90, HER2, and urease. In addition, the cultivation of _A. ficuum_ extracts in a controlled environment with the progress of new approaches will lead to the discovery of new
bioactive chemical scaffolds in the drug discovery process. DATA AVAILABILITY All data generated or analyzed during this study are included in this published article and its supplementary
information files. REFERENCES * Hoeksma, J. _et al._ A new perspective on fungal metabolites: identification of bioactive compounds from fungi using zebrafish embryogenesis as read-out.
_Sci. Rep._ 9, 17546 (2019). PubMed PubMed Central ADS Google Scholar * Elkhouly, H. I. _et al._ Bioactive secondary metabolite from endophytic Aspergillus tubenginses ASH4 isolated from
Hyoscyamus muticus: Antimicrobial, antibiofilm, antioxidant and anticancer activity. _Pharmacogn. J._ 13(2), 434–442 (2021). CAS Google Scholar * Zheng, R. _et al._ Biological activities
of some new secondary metabolites isolated from endophytic fungi: A review study. _Int. J. Mol. Sci._ 22(2), 959 (2021). MathSciNet CAS PubMed PubMed Central Google Scholar * Devi, R.
_et al._ Fungal secondary metabolites and their biotechnological applications for human health. In _New and Future Developments in Microbial Biotechnology and Bioengineering_ (eds Asghar, A.
_et al._) 147–161 (Elsevier, 2020). Google Scholar * Hou, X. _et al._ Recent advances in sorbicillinoids from fungi and their bioactivities. _J. Fungi_ 8(1), 62 (2022). MathSciNet CAS
Google Scholar * Elshafei, A. M. & El-Ghonemy, D. H. Screening and media optimization for enhancing L-asparaginase production, an anticancer agent, from different filamentous fungi in
solid state fermentation. _Br. Biotechnol. J._ 9(3), 1–15 (2015). Google Scholar * Qi, C. _et al._ Butenolides from a marine-derived fungus Aspergillus terreus with antitumor activities
against pancreatic ductal adenocarcinoma cells. _Bioorg. Med. Chem._ 26(22), 5903–5910 (2018). CAS PubMed Google Scholar * Nguyen, V.-T. _et al._ Gliotoxin isolated from marine fungus
Aspergillus sp. induces apoptosis of human cervical cancer and chondrosarcoma cells. _Mar. Drugs_ 12(1), 69–87 (2013). PubMed PubMed Central Google Scholar * Ghfar, A. A. _et al._
Production of terretonin N and butyrolactone I by thermophilic aspergillus terreus TM8 promoted apoptosis and cell death in human prostate and ovarian cancer cells. _Molecules_ 26(9), 2816
(2021). CAS PubMed PubMed Central Google Scholar * Taritla, S. _et al._ Optimization of physicochemical parameters for production of cytotoxic secondary metabolites and apoptosis
induction activities in the culture extract of a marine algal-derived endophytic fungus Aspergillus sp. _Front. Pharmacol._ 12, 542891 (2021). CAS PubMed PubMed Central Google Scholar *
Mazumder, K. _et al._ Identification of bioactive metabolites and evaluation of in vitro anti-inflammatory and in vivo antinociceptive and antiarthritic activities of endophyte fungi
isolated from Elaeocarpus floribundus blume. _J. Ethnopharmacol._ 273, 113975 (2021). CAS PubMed Google Scholar * Nielsen, K. F. _et al._ Review of secondary metabolites and mycotoxins
from the Aspergillus niger group. _Anal. Bioanal. Chem._ 395, 1225–1242 (2009). CAS PubMed Google Scholar * Sun, J. _et al._ Metabolic peculiarities of Aspergillus niger disclosed by
comparative metabolic genomics. _Genome Biol._ 8(9), R182 (2007). PubMed PubMed Central Google Scholar * Shah, Z. A. _et al._ Insights into metabolic and pharmacological profiling of
Aspergillus ficuum through bioinformatics and experimental techniques. _BMC Microbiol._ 22, 295 (2022). CAS PubMed PubMed Central Google Scholar * National Research Council. _Guide for
the Care and Use of Laboratory Animals_ 8th edn. (The National Academies Press, 2011). Google Scholar * Velez, M. E. V. _et al._ Production of bioactive compounds by the mycelial growth of
Pleurotus djamor in whey powder enriched with selenium. _LWT-Food Sci. Technol._ 114, 108376 (2019). CAS Google Scholar * Shah, Z. A. _et al._ Metabolic and pharmacological profiling of
Penicillium claviforme by a combination of experimental and bioinformatic approaches. _Ann. Med._ 54(1), 2102–2114 (2022). Google Scholar * Kamble, R. _et al._ Evaluation of antispasmodic
activity of different Shodhit guggul using different shodhan process. _Indian J. Pharm. Sci._ 70(3), 368–372 (2008). PubMed PubMed Central Google Scholar * Sisay, M. _et al._ Evaluation
of the antispasmodic and antisecretory activities of the 80% methanol extracts of verbena officinalis L: Evidence from in vivo antidiarrheal study. _J. Evid. Based Integr. Med._ 24,
2515690X19853264 (2019). CAS PubMed PubMed Central Google Scholar * Florento, L. _et al._ Comparison of cytotoxic activity of anticancer drugs against various human tumor cell lines
using in vitro cell-based approach. _Int. J. Biomed. Sci._ 8(1), 76–80 (2012). CAS PubMed PubMed Central Google Scholar * Hanif, M. _et al._ Synthesis, urease inhibition, antioxidant,
antibacterial, and molecular docking studies of 1, 3, 4-oxadiazole derivatives. _ISRN Pharmacol._ 2012, 1–9 (2012). ADS Google Scholar * Balasubramanian, A. & Ponnuraj, K. Crystal
structure of the first plant urease from jack bean: 83 years of journey from its first crystal to molecular structure. _J. Mol. Biol._ 400(3), 274–283 (2010). CAS PubMed Google Scholar *
Aertgeerts, K. _et al._ Structural analysis of the mechanism of inhibition and allosteric activation of the kinase domain of HER2 protein. _J. Biol. Chem._ 286(21), 18756–18765 (2011). CAS
PubMed PubMed Central Google Scholar * Molecular Operating Environment (MOE), 2011. 10; Chemical Computing Group Inc., Montreal, QC, Canada, 2012. * Jia, C. _et al._ Antifungal activity
of coumarin against Candida albicans is related to apoptosis. _Front. Cell. Infect. Microbiol._ 8, 1–13 (2019). Google Scholar * Tran, H. N. _et al._ In vitro antifungal activity of
Cinnamomum zeylanicum bark and leaf essential oils against Candida albicans and Candida auris. _Appl. Microbiol. Biotechnol._ 104(20), 8911–8924 (2020). CAS PubMed PubMed Central Google
Scholar * Bladt, T. T. _et al._ Anticancer and antifungal compounds from Aspergillus, Penicillium and other filamentous fungi. _Molecules_ 18(9), 11338–11376 (2013). CAS PubMed PubMed
Central Google Scholar * Svahn, K. S. _et al._ Antimicrobial activity of filamentous fungi isolated from highly antibiotic-contaminated river sediment. _Infect. Ecol. Epidemiol._ 2, 11591
(2012). Google Scholar * Dreyfuss, M. & Chapela, I. H. Potential of fungi in the discovery of novel, low-molecular weight pharmaceuticals. In _Discovery of Novel Natural Products with
Therapeutic Potential_ (ed. Vincent, P. G.) 49–80 (Elsevier, 1994). Google Scholar * Bugni, T. S. _et al._ Yanuthones: Novel metabolites from a marine isolate ofAspergillus niger. _J. Org.
Chem._ 65(21), 7195–7200 (2000). CAS PubMed Google Scholar * Teixeira, M. F. _et al._ Amazonian biodiversity: pigments from Aspergillus and Penicillium-characterizations, antibacterial
activities and their toxicities. _Curr. Trends Biotechnol. Pharm._ 6(3), 300–311 (2012). CAS Google Scholar * Vargas, R. _et al._ Relaxant and antispasmodic effect in isolated guinea pig
ileum treated with extracts of Xylaria sp an endophytic fungus of the Mexican yew, Taxus globosa. _Pharmacologyonline_ 2, 134–145 (2007). Google Scholar * Severiano, M. E. _et al._
Biotransformation of ent-pimaradienoic acid by cell cultures of Aspergillus niger. _Bioorg. Med. Chem._ 21(18), 5870–5875 (2013). CAS PubMed Google Scholar * Kharwar, R. N. _et al._
Anticancer compounds derived from fungal endophytes: Their importance and future challenges. _Nat. Prod. Rep._ 28, 1208–1228 (2011). CAS PubMed Google Scholar * Minarni _et al._
Anticancer activity test of ethyl acetate extract of endophytic fungi isolated from soursop leaf (Annona muricata L.). _Asian Pac. J. Trop. Med_. 10(6), 566–571 (2017). * Felczykowska, A.
_et al._ Antibacterial and anticancer activities of acetone extracts from in vitro cultured lichen-forming fungi. _BMC Complement Altern. Med._ 17(1), 1–12 (2017). Google Scholar *
Majoumouo, M. S. _et al._ Cytotoxicity potential of endophytic fungi extracts from Terminalia catappa against human cervical cancer cells. _J. Toxicol._ 2020, 1–9 (2020). Google Scholar *
Thomas, A. T. _et al._ In vitro anticancer activity of microbial isolates from diverse habitats. _Braz. J. Pharm. Sci._ 47(2), 279–287 (2011). CAS Google Scholar * Nadumane, V. K.,
Venkatachalam, P. & Gajaraj, B. Aspergillus applications in cancer research. In _New and Future Developments in Microbial Biotechnology and Bioengineering_ (ed. Gupta, V. K.) 243–255
(Elsevier, 2016). Google Scholar * Miao, L. _et al._ Effect of culture conditions on mycelial growth, antibacterial activity, and metabolite profiles of the marine-derived fungus Arthrinium
cf saccharicola. _Appl. Microbiol. Biotechnol._ 72(5), 1063–1073 (2006). CAS PubMed Google Scholar * Bilal, S. _et al._ Endophytic fungus Paecilomyces formosus LHL10 produces
sester-terpenoid YW3548 and cyclic peptide that inhibit urease and α-glucosidase enzyme activities. _Arch. Microbiol._ 200(10), 1493–1502 (2018). CAS PubMed Google Scholar * Rauf, A. _et
al._ Urease inhibition potential of Di-naphthodiospyrol from Diospyros lotus roots. _Nat. Prod. Res._ 31(10), 1214–1218 (2017). CAS PubMed ADS Google Scholar * Doriya, K. & Kumar, D.
S. Isolation and screening of L-asparaginase free of glutaminase and urease from fungal sp. _3 Biotech._ 6, 239 (2016). PubMed PubMed Central Google Scholar * Baazeem, A. _et al._
Paecilomyces formosus MD12, a biocontrol agent to treat Meloidogyne incognita on brinjal in green house. _J. Fungi (Basel)_ 7(8), 632 (2021). CAS PubMed Google Scholar * Khan, A. L. _et
al._ Enzyme inhibitory radicinol derivative from endophytic fungus Bipolaris sorokiniana LK12, associated with Rhazya stricta. _Molecules_ 20(7), 12198–12208 (2015). CAS PubMed PubMed
Central Google Scholar * Matthews, R. & Burnie, J. The role of hsp90 in fungal infection. _Immunol. Today_ 13(9), 345–348 (1992). CAS PubMed Google Scholar * Gutierrez, C. &
Schiff, R. HER2: Biology, detection, and clinical implications. _Arch. Pathol. Lab. Med._ 135(1), 55–62 (2011). PubMed PubMed Central Google Scholar * Belhassan, A. _et al._ In silico
detection of potential inhibitors from vitamins and their derivatives compounds against SARS-CoV-2 main protease by using molecular docking, molecular dynamic simulation and ADMET profiling.
_J. Mol. Struct._ 1258, 132652 (2022). CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS The authors are grateful to the Department of Chemistry, Islamia
College, Peshawar, KP, Pakistan for the provision of all facilities needed to execute the project. The authors are also thankful to Universidade Federal de Pelotas—UFPel, Pelota, RS, Brazil
for providing support to perform this project. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Chemistry, Islamia College University, Peshawar, Khyber Pakhtunkhwa, Pakistan Zafar
Ali Shah, Khalid Khan, Nasir Ahmad & Akhtar Muhammad * Institute of Basic Medical Sciences, Khyber Medical University, Peshawar, Khyber Pakhtunkhwa, Pakistan Tanzeel Shah * Center of
Chemical, Pharmaceutical and Food Sciences, Federal University of Pelotas, Pelotas RS, Brazil Haroon ur Rashid * Institute of Chemistry, Sao Paulo State University, Araraquara, Sao Paulo,
Brazil Haroon ur Rashid Authors * Zafar Ali Shah View author publications You can also search for this author inPubMed Google Scholar * Khalid Khan View author publications You can also
search for this author inPubMed Google Scholar * Tanzeel Shah View author publications You can also search for this author inPubMed Google Scholar * Nasir Ahmad View author publications You
can also search for this author inPubMed Google Scholar * Akhtar Muhammad View author publications You can also search for this author inPubMed Google Scholar * Haroon ur Rashid View author
publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS Z.A.S.: conceptualization, formal analysis, investigation, and methodology; K.K.: conceptualization,
supervision, project administration, data curation, and resources; T.S.: formal analysis, validation, and writing—review, and editing; N.A.: formal analysis and software; A.M.: formal
analysis, methodology, and writing—review and editing; H.R.: data curation, software, and writing—review, and editing. All authors agree to publish the data. CORRESPONDING AUTHORS
Correspondence to Khalid Khan or Haroon ur Rashid. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer
Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN
ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or
other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not
included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission
directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Shah,
Z.A., Khan, K., Shah, T. _et al._ Biological investigations of _Aspergillus ficuum_ via in vivo_, _in vitro and in silico analyses. _Sci Rep_ 13, 17260 (2023).
https://doi.org/10.1038/s41598-023-43819-y Download citation * Received: 03 May 2023 * Accepted: 28 September 2023 * Published: 11 October 2023 * DOI:
https://doi.org/10.1038/s41598-023-43819-y SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative