Play all audios:
ABSTRACT Dissolved inorganic nutrient concentrations in the surface waters (0 to 5 m) of the Northern Gulf of Mexico (NGoM) were analyzed from 1985 to 2019 (> 10,000 observations) to
determine spatiotemporal trends and their connection to nutrients supplied from the Mississippi/Atchafalaya River (MAR). In the NGoM, annual mean dissolved inorganic P (DIP) concentrations
increased significantly over time, while dissolved inorganic N (DIN) concentrations showed no temporal trend. With greater salinity, mean DIN:DIP decreased from above the Redfield ratio of
16 to below it, reflecting DIN losses and the more conservative behavior of DIP with salinity. Over the same time period, annual mean P (total dissolved P, DIP, dissolved organic P) loading
from the MAR to the NGoM significantly increased, annual mean DIN and total dissolved N loading showed no temporal trend, and dissolved organic N loading significantly decreased. Though DIP
increased in the MAR, MAR DIP alone was insufficient to explain the surface distribution of DIP with salinity. Therefore, increases in surface DIP in the NGoM are not simply a reflection of
increasing MAR DIP, pointing to temporal changes in other DIP sources. The increase in NGoM DIP suggests greater N limitation for phytoplankton, with implications for N fixation and nutrient
management. SIMILAR CONTENT BEING VIEWED BY OTHERS ATMOSPHERIC DEPOSITION AND RIVER RUNOFF STIMULATE THE UTILIZATION OF DISSOLVED ORGANIC PHOSPHORUS IN COASTAL SEAS Article Open access 30
January 2024 INTERMITTENT MEROMIXIS CONTROLS THE TROPHIC STATE OF WARMING DEEP LAKES Article Open access 31 July 2020 NUTRIENT LOADING AS A KEY CAUSE OF SHORT- AND LONG-TERM ANTHROPOGENIC
ECOLOGICAL DEGRADATION OF THE SALTON SEA Article Open access 28 December 2024 INTRODUCTION Marine primary production is often mediated and limited by the bioavailability of dissolved
nutrients such as nitrogen (N) and phosphorus (P)1,2,3. Studies have shown that N limitation of marine primary production is more widespread than P limitation3,4, though P availability may
play an important role over long time scales5,6, and in certain locations, such as the Northern Gulf of Mexico7. Surface ocean N and P concentrations are spatially and temporally variable as
a result of many complex processes such as uptake by phytoplankton and bacteria, including luxury consumption8, N fixation9, N loss through denitrification in low oxygen regions10,
biological and chemical conversion of inorganic and organic N and P11,12, legacy nutrients stored in the landscape13,14, and external anthropogenic inputs15,16. These processes can lead to
deviations in organic matter production and dissolved nutrient ratios from the canonical 106C:16N:1P of Redfield proportions3,12,17. For example, rivers, estuaries, and coastal regions
typically have higher dissolved inorganic N and P (DIN and DIP) concentrations and DIN:DIP than offshore regions18, where average surface DIN:DIP is about 135. Meanwhile, in the majority (~
78%) of the world’s large rivers, DIN:DIP exceeds 16, and increases with DIN concentrations19. These spatial patterns could be explained by net relative losses of DIN5,20, and/or by a
relative net gain of DIP from bioconversion of dissolved organic P (DOP) to DIP as salinity increases from nearshore to offshore waters21,22. In this study, we focus on the spatial and
temporal variability of surface DIN and DIP concentrations over the last 35 years in the Northern Gulf of Mexico (NGoM) along the salinity gradient from the Mississippi and Atchafalaya
Rivers (MAR) to offshore oceanic waters. Annual mean MAR discharge is more than 15 times that of all other rivers that drain into the NGoM23, thus we focus on annual mean MAR discharge as
the main source of nutrients and freshwater into the NGoM system. The size, direction, and location of the NGoM freshwater plume change in concert with varying volume and timing of river
discharge as well as wind speed and direction, therefore further influencing nutrient and salinity patterns24,25,26. Riverine freshwater plumes generally extend westward in the NGoM through
the Louisiana Coastal Current, though wind forcing pushes buoyant plumes eastward depending on the time of year23,27. Physical drivers such as onshore winds and salinity cause MAR plume
waters and its nutrients, sediments, and organic matter to be transported westward alongshore and eastward along the approximately 200 m depth shelf break27,28. The majority of riverine N
and P are retained in nearshore regions of the NGoM in the fall and winter29, and spread offshore in the summer, though N typically declines more dramatically as a function of salinity than
P30,31. When averaged annually, the majority of surface water DIN (70%) is retained on the shelf, while 30% of DIN is transported further offshore32. Together the MAR are the main sources of
freshwater and nutrients into the NGoM, on average delivering 80% of the freshwater, 91% of the N loading, and 88% of the P loading into the system with a combined mean flow of
approximately 21,500 m3 s−133,34,35. Over the last 200 years, many aspects of the MAR watershed have been altered by changing water demands, fluctuating sediment yields, navigational
amendments, and flood-control systems36. The MAR’s water quality and chemistry has been substantially impacted by changes in land use, agriculture, industry, and sewage effluent37,38. From
the 1950s to 1990s, TDN loading (primarily driven by increasing DIN loading) from the MAR to the NGoM tripled, and TDP loading doubled35,36. Since then, TDN loading has not appreciably
increased, and has even stabilized in some locations35,39. Earlier studies found no temporal trends in DIP or TDP from the 1970s to 1990s40. Temporal trends in MAR nutrient loading are
similar to global trends in the latter part of the twentieth century, though MAR N fluxes increased more, and P fluxes increased less, than the global average41. Despite the increase in N
loading from the MAR, empirical studies indicate a predominance of N limitation of phytoplankton in the NGoM11, and isotopic evidence indicates that the majority of N incorporated into
planktonic biomass in the NGoM originates from MAR loading35. Nevertheless, observations of P limitation have been reported, especially at intermediate salinities within the MAR plume during
spring and summer42,43,44. Multiple studies have investigated the connections between MAR flow and NGoM nutrient concentrations31,45,46,47. Lohrenz et al. (1999) found a positive
correlation between MAR river discharge and MAR N:P, and in their 1990 study concluded that riverine nutrient supply constraints were a controlling factor of biomass and production at high
salinities. Wysocki et al. (2006) further established that the spatial distribution of NGoM nutrients changed with MAR flow, with higher NGoM nutrient concentrations observed further
offshore during periods of higher discharge. However, Cardona et al. (2016) concluded that MAR discharge alone was insufficient to predict NGoM surface nutrient concentrations, given low
nutrient concentrations observed following high flow periods. Additionally, MAR discharge and nutrient flux are tied to the spatial and temporal variability of the summer hypoxic area, or
“dead zone” in the NGoM (characterized by dissolved oxygen content of < 2 mg L−1)48. The increase in MAR DIN loading from the 1950s to 1990s coincided with increased NGoM primary
production, sediment C accumulation, and hypoxia—hypoxia did not appear as widespread or recurrent prior to the 1950s increase in DIN35,49,50. The areal extent of the NGoM dead zone is also
correlated with MAR DIN loading and with primary production only in the MAR plume, not the full shelf area of the GoM35,44,51. Over time, the relationship between DIN loading and hypoxic
area has changed, with the same amount of DIN loading in recent years leading to larger hypoxic areas than prior to the early 1990s52,53. Despite the importance of DIN in NGoM hypoxia, model
simulations show that P limitation may play an important role by shifting primary production downstream within the plume and decreasing the area of hypoxic bottom water, due to changes in
where primary production occurs and whether respiration occurs in the sediment or water column54. Field studies are consistent with model findings, as P limitation of phytoplankton delays
the assimilation of riverine DIN in the summer and drives primary production over a larger region beyond shelf plume waters44. The purpose of this study was to understand the patterns of
DIN, DIP, and DIN:DIP on multiple scales in the NGoM; temporally (1985 to 2019) and spatially (shelf to offshore) in surface waters (0 to 5 m). This study represents an expanded view of
nutrient trends and spatial patterns in the NGoM in the context of ongoing efforts to manage MAR nutrient loads to the NGoM, especially for N55. Many other studies have characterized the
surface nutrient trends in the NGoM, though more have focused on more limited areas or timespans31,36,56. Based on our temporal and spatial nutrient analyses, we address the following
objectives: (a) to delineate spatiotemporal trends in surface nutrient (DIN and DIP) concentrations and in the resulting DIN:DIP in the MAR and NGoM from 1985 to 2019; (b) to evaluate MAR
nutrients as potential drivers of NGoM nutrient change over time and space; and (c) to determine whether nutrient shifts described in previous studies have persisted (i.e., changes in
nutrient regime, anthropogenic impacts). These objectives aim to examine how changes in nutrient delivery through the MAR contribute to variations in surface nutrients in the NGoM over time
and space. METHODOLOGY DATA DESCRIPTION The NGoM nutrient data set was compiled following methods used in Cardona et al. (2016), and enhanced by including a larger salinity range (i.e.,
including salinities lower than 11 ppt), adding additional nutrient samples post-2012, and other pre-2012 observations that had not been included in the earlier study. Our study’s data set
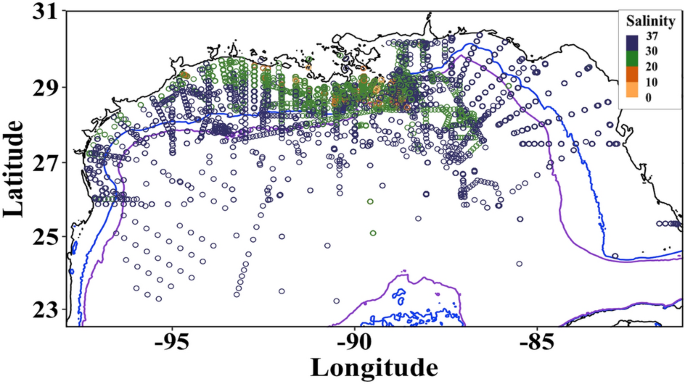
included only surface (0 to 5 m collection depth) nutrient data in the NGoM (defined as coordinates − 98˚, − 79˚ to 22.5˚, 31˚; Fig. 1) from 1985 to 2019, with most data collected in the
summer months (Fig. S2). NGoM surface data were used to ensure that samples were influenced by the MAR plume. Numerous studies of nutrients, phytoplankton, and riverine transport have
entirely or primarily focused on the upper few meters of the NGoM water column (e.g., Cardona et al., 2016, Wysocki et al., 2006). Thus, the results of this study elaborate on the context
provided by earlier work. The criteria for the compiled NGoM surface nutrient data included: 0 to 5 m sampling depth, collection date from 1985 to 2019, and study location in the northern
portion of the GoM (within coordinate box listed above). Data also needed to include salinity (0–37 ppt), and surface DIN and DIP concentrations in µM, if data were not listed in µmoles/L or
µM, they were converted to µM. For quality assurance, data with salinities greater than 37 ppt or without corresponding salinity values were excluded (i.e., this excluded all data points
from cruises from 1988 and 2014). In addition, 39 NGoM datapoints (from n = 10,007 total) with nutrient values above 8 µM DIP or 110 µM DIN were excluded as they were greater than three
standard deviations above the mean. The bottom depth for each sample coordinate was calculated with the _marmap_ R package for all data within coordinates − 98°, − 79°, and 20°, 31°57. These
data were compiled from a variety of sources with varied collection and analysis data validation methods; with this in mind, we verified the data in comparison to data from other years and
regions, covering a large portion of the surface waters of the NGoM over 35 years. Data with these criteria were compiled from the Biological & Chemical Oceanography Data Management
Office (BCO-DMO), National Oceanic and Atmospheric Administration National Centers for Environmental Information—World Ocean Database (NOAA NCEI WOD), United States Geological Survey (USGS),
and Gulf of Mexico Research Initiative Information and Data Cooperative (GRIIDC) (Table S1). Uptake rates of NH4+ in the MAR plume can be comparable to those for NO3-; however, we
operationally defined DIN as the NO2− + NO3− concentration, excluding NH4+ because it was relatively rarely measured in the MAR and NGoM databases and was often near or below the detection
limit or a minor fraction of DIN when it was quantified. Typically, NH4+ is rapidly transformed into NO2− + NO3−, with concentrations from 0.17 to 0.44 µM and little spatial variability in
both the lower MAR and NGoM58,59. Prior studies of this region similarly focused on NO2− + NO3−. USGS historical records for the Mississippi River at St. Francisville, LA, that we used in
our MAR nutrient comparison31,60,61, also define inorganic N as NO2− + NO3−. The majority of surveyed studies did not measure organic forms of N and P, so analyses were conducted only using
inorganic nutrient data. For some analyses, the surface nutrient data were subset into three different spatial regions: hypoxic region (data within − 88° to − 95° and 27° to 29° with bottom
depths of < 60 m, Figure S3a), shelf region (data with bottom depths of < 200 m, Fig. S3b), and offshore region (data with bottom depths of > 200 m, Fig. S3c). The hypoxic region
defined here was a shallow subset of the shelf region where hypoxic bottom waters are most likely to be found; the boundaries of the hypoxic region did not change through time in these
analyses, though the actual area measured with hypoxic bottom water varies seasonally and annually35. MAR data (nutrient loading, nutrient concentrations, discharge, and suspended sediment
concentrations) were also compiled for 1985 to 2019 from the St. Francisville, Louisiana USGS National Water Quality Network (NWQN) program monitoring station because it had the longest
running nutrient data (dissolved nutrients measured multiple times per year since 1954) in the lower portion of the MAR, and it is located close to the point where the Atchafalaya River
diverges from the Mississippi, covering a drainage area greater than 2.9 × 106 km262. Many other NGoM studies similarly use MAR nutrient data from the St. Francisville, LA USGS monitoring
station36,63,64. In addition to DIN (NO2− + NO3− as N, filtered, mg L−1) and DIP (orthophosphate, filtered, mg L−1), the USGS MAR water quality data included TDN (NO2− + NO3− + NH4+ +
organic-N, filtered, mg L−1), TDP (filtered, mg L−1), dissolved organic N (DON; filtered, mg L−1), and dissolved organic P (DOP; filtered, mg L−1), that were not included within the NGoM
nutrient data set because they were rarely measured compared to DIN and DIP in the NGoM. MAR data were compiled for nutrients (in mg L−1), discharge (in tons, then converted to kg year−1),
loading (in tons, then converted to kg year−1) and suspended sediments from all available dates between 1985 and 2019. The MAR basin has been a location of intense streamflow and large-scale
water quality monitoring for decades; this study tests whether the MAR nutrient fluxes can explain spatial and temporal nutrient trends in the NGoM as a whole. MAR St. Francisville water
quality loads are sourced from USGS (USGS Station 07373420) as indicated in Lee (2022). USGS computed annual, flow-normalized water-quality loads (mean annual sample n = 16) using the USGS
Load Estimator (LOADEST) program and Weighted Regressions on Time, Discharge, and Season method (WRTDS) between 1985 and 2019 using available discrete water-quality and streamflow
information61. Nutrient fluxes were calculated using Adjusted Maximum Likelihood Estimation (AMLE) using the LOADEST program to compute nutrient loads using a 10-year moving window approach
in the MAR basin61,65. Load estimation methods included the log of cubic streamflow, time (annual, seasonal, monthly), and historical streamflow conditions61. The WRTDS water quality data
for the MAR is used by the Mississippi River/Gulf of Mexico Hypoxia Task Force to meet their goals of reducing the hypoxic zone in the GoM to a 5 year moving average of 5000 km261,65. To
determine the annual amount of MAR discharge at St. Francisville, annual flow for a given water year was converted from daily cubic meters second−1 to acre-feet day−1 then averaged annually.
For WRTDS loads calculated for the MAR at St, Francisville, WRTDS calibration records existed from 1980 to 2019 for loads of TDN, TDP, DIN, and suspended sediment concentrations, while DIP
loads were analyzed and calibrated from 1982 to 201961,65. These WRTDS loads are assumed to be the most accurate load estimates for any given year65. In our study, the MAR annual WRTDS
nutrient load data was compared to MAR nutrient data using comparable analyses to determine correlations and compare trends in nutrient concentrations over time and space. DATA ANALYSES All
statistical analyses used a significance level of 0.05. NGoM temporal nutrient trends were evaluated by linear regressions of annual mean DIN and DIP concentrations and DIN:DIP against time.
Similarly, temporal trends in MAR data were assessed using linear regressions of annual means for each parameter over time (nutrient loading, nutrient concentrations, discharge, suspended
sediment). Temporal regression analyses in this study set 1985 as year zero so that the regression equations provided meaningful y-intercepts. Oftentimes, long successive time-series contain
autocorrelation of data66. To remove potential autocorrelation in the time series data, we based the analysis on annual mean NGoM and MAR nutrient concentrations, and also tested for
autocorrelation using Durbin-Watson tests. For data with significant autocorrelation, Cochrane-Orcutt transformation was used, and Durbin-Watson tests were run again to confirm reduction of
autocorrelation below significance. The Cochrane-Orcutt estimation and subsequent transformation also accounted for heteroscedasticity in the data, confirmed by Breusch-Pagan tests. In
addition to linear regression, changepoint analyses of mean annual nutrient concentrations were conducted using a regression model in R package _changepoint_ with segmented relationships for
annual mean DIN and DIP over time to determine whether monotonic analyses were appropriate for the nutrient time series. Changepoint analyses identify statistically significant changes or
breaks in trends over time. After standardizing year and nutrient concentration (DIN, DIP) variables, we then used a Markov Chain Monte Carlo simulation to fit a Bayesian changepoint model.
The changepoint in the data represents a gap or change in the distribution of the nutrient data in a given year. If there are no significant changepoints, then monotonic analyses are best
suited for the data. Pearson correlation analyses tested for significant temporal relationships between annual mean MAR discharge, nutrient concentrations, and nutrient loading with the
corresponding annual mean NGoM nutrient concentrations. Parallel analyses focused on each of the three regional data subsets: hypoxic, shelf, and offshore regions. Pearson correlation
analyses also compared annual mean nutrient concentrations in the shelf and offshore regions to each other. Finally, Pearson correlation analyses compared the annual area of hypoxic bottom
water35 to mean annual MAR nutrient loading and mean annual NGoM nutrient concentrations as a whole, as well as in each spatial region. Next, the NGoM nutrient data set was analyzed in
relation to sample salinity to incorporate dilution along the continuum from the MAR endmember to the oceanic endmember into the analyses of nutrient concentrations. While there was
generally a trend of increasing salinity going offshore, it should be noted that nearly the full range of salinities was found in both shelf and offshore regions, and salinity plots were
thus not strictly analogous to geographic patterns (i.e., some samples found beyond the 200 m isobath had salinities less than 20; Fig. S2). Low salinity offshore waters were located within
the freshwater plume, which varies over time and space with environmental variables such as river discharge, wind speed, and direction67. Nutrient data were regressed against salinity using
a variety of functions, including linear, exponential, and power functions; within monotonic functions, bivariate linear regression of ln-transformed data provided the highest r2 values and
significance. To account for zero values in the data set prior to the ln-transformation, we added a reasonable detection limit to all samples (0.05 µM for DIN, 0.03 µM for DIP), similar to
that used in Cardona et al. (2016). Linear regressions of ln[DIN], ln[DIP], and ln(DIN:DIP) vs. salinity described the general trends of decreasing nutrient concentrations along the salinity
gradient due to dilution and loss (e.g., phytoplankton uptake). To improve visualization, especially of low nutrient concentrations, figures show untransformed data plotted with log-scale
y-axes. These regressions and their presentation on plots were primarily intended to show general trends of nutrient concentrations with salinity. Nutrient data were also compared to
conservative mixing functions, calculated by linearly connecting the long-term mean MAR TDN, TDP, DIN, and DIP concentrations at St. Francisville, LA at 0 ppt salinity to the minimum ocean
endmember nutrient concentration (the detection limits mentioned above) at 37 ppt (Fig. S1). This line described the expected decline of nutrient concentration due to dilution of MAR water
with the offshore endmember. Note that on the semi-log plots shown in later figures, the mixing functions appear curved. To understand how the relationship between nutrient concentrations
and salinity changed through time relative to the conservative mixing function, we calculated the residuals of each data point compared to the respective conservative mixing function along
the salinity gradient. Negative residuals signified loss of nutrients relative to the MAR endmember that exceeded the decline expected due to mixing with the offshore endmember (e.g., due to
phytoplankton uptake). Positive residuals indicated excess inorganic nutrients relative to the two-endmember function, suggesting a source in addition to the MAR endmember. Analogous
residual analyses were also conducted on the hypoxic, shelf, and offshore regional subsets. RESULTS SPATIOTEMPORAL CHANGES IN ANNUAL MEAN NGOM DIN, DIP, AND DIN:DIP NGoM DIN concentrations
(1985–2019) were highly variable and showed no significant temporal trend (Fig. 2a), while DIP concentrations increased significantly (Fig. 2b). DIN:DIP also showed no significant temporal
trend, though DIN:DIP ratios from 2015 to 2019 were among the lowest of the study period, with a mean DIN:DIP of 3.3 (Fig. 2c). Low DIN:DIP values (from 1.2 to 3.6) also occurred in the late
1980s. Durbin-Watson autocorrelation tests of annual mean nutrient concentrations over time showed no autocorrelation for annual DIN means over time, and minimal autocorrelation for annual
DIP means over time (Table S4). Thus, Cochrane-Orcutt estimations were run on the annual DIP means to transform the data and remove autocorrelation, then Durbin-Watson autocorrelation tests
were run again to confirm there was no longer significant autocorrelation. The transformed DIP data had the same r2 value, _p_-value, and slope as the untransformed data, and therefore the
untransformed data is shown in Fig. 2 for clarity. In addition, the transformation removed heteroscedasticity in the DIP means over time, which was confirmed with the Breusch-Pagan test.
Bayesian changepoint analyses of DIN and DIP over time showed that there were no years when there was a significant changepoint in the time series; though there was a decrease in the
changepoint time statistic for DIN at 2013 and an increase in the changepoint time statistic for DIP at 1999, the error bars for each were large and included much of the time series (Fig.
S5). There were also no significant temporal DIN or DIN:DIP trends in the regional subsets; however, hypoxic and shelf DIP concentrations significantly increased over time (Fig. S6; Table
S3). ANNUAL MEAN MAR NUTRIENT LOADING INTO THE NGOM Similar to NGoM nutrient trends from 1985 to 2019, TDN and DIN loading from the MAR into the NGoM did not change significantly, while DON
loading significantly decreased (Fig. 3a, Table S2). Over the same time period, TDP, DIP, and DOP loading from the MAR into the NGoM all increased significantly (Fig. 3b, Table S2). As
annual mean MAR discharge did not change significantly over the study time period (Fig. S4b), it is not surprising that MAR DIN concentrations did not show any temporal trend from 1985 to
2019, although MAR DIP concentrations increased significantly (Fig. S4a). Annual mean suspended sediment concentrations, however, declined significantly during the period (Fig. S4c).
Comparing annual mean NGoM DIN and DIP concentrations to MAR discharge, MAR DIN and DIP concentrations, and MAR DIN and DIP loads produced similar patterns of significant relationships for
the entire NGoM, the hypoxic region, and the shelf region (Table S4). All were significantly correlated except DIN:DIP, which did not significantly correlate to MAR discharge in the entire
NGoM, hypoxic, or shelf regions. In contrast to results for the hypoxic and shelf regions, correlation analyses for the offshore region found no significant correlations between offshore DIN
and MAR discharge, MAR DIN or DIP concentrations or loading (Table S4). On the other hand, offshore DIP concentrations did significantly correlate with MAR discharge, MAR DIP
concentrations, and MAR DIN:DIP. Offshore DIN:DIP was significantly correlated to MAR discharge, MAR DIN and DIP concentrations and loading, however, not to MAR DIN:DIP (Table S4). In
general, shelf and offshore nutrient concentrations were not significantly correlated with each other over time, with exceptions of the significantly positively correlated offshore DIN:DIP
to shelf DIN and DIP concentrations (Table S5). The areal extent of the hypoxic bottom water in the NGoM fluctuates interannually35, and correlation analyses were used to determine if this
hypoxic area related to nutrient observations. The annual area of hypoxic bottom water was significantly correlated with all forms of N and P loading from the MAR into the NGoM (Table S6).
The annual area of hypoxic bottom water was also significantly correlated with annual mean NGoM DIN and DIP concentrations and DIN:DIP in the entire NGoM dataset, the hypoxic region, and the
shelf region (Table S6). However, the area of hypoxic bottom water was not significantly correlated with offshore nutrient concentrations (Table S6). NUTRIENT VERSUS SALINITY RELATIONSHIPS
IN THE NGOM All NGoM DIN and DIP data (1985–2019) were plotted against corresponding sample salinity in Fig. 4 with each black circle representing a single sample. The conservative mixing
function and linear best fits (to ln-transformed values) were also shown. Both DIN and DIP generally declined with increasing salinity, and the slope of the decline was steeper for DIN
compared to DIP. Relative to the respective conservative mixing function (shown in purple), only 2% of the DIN values across the salinity range were above the DIN function, while about half
(51.8%) of the DIP observations exceeded the DIP mixing function. DIN:DIP in the NGoM generally decreased from above to below the Redfield ratio with increasing salinity, driven by the
greater decline in DIN relative to DIP concentrations (Fig. 4c). At 0 salinity in the MAR, DIN:DIP averaged 46.8, and typically declined to less than 16 within the NGoM where salinity was
greater than 20. Nutrient vs. salinity relationships were also plotted separately for the hypoxic, shelf, and offshore subsets of the data (Fig. S7; Table S6). While there were differences
in the best-fit slopes for each of the three data subsets, the general patterns were similar to the overall data set. Changing the freshwater endmember across the range of DIN and DIP values
from 1985 to 2019 did not affect nutrient vs. salinity relationships. For all regions, DIN declined more for a given change in salinity than DIP, and a much larger fraction of the DIP
observations (relative to the DIN observations) exceeded the mixing function. Subsequent analyses subset the data shown in Fig. 4 to examine the residuals of each nutrient from the mixing
functions. Nutrient residuals relative to the conservative mixing function were calculated for each data point as the difference between actual and predicted value, with positive residuals
meaning an excess or production of the nutrient while the negative residuals indicate the loss or consumption of the nutrient in addition to dilution by low-nutrient ocean water (depending
on the sample salinity; Fig. 5, Figs. S8, S9). Here, we emphasize the presentation of data on the DIP residuals because over half exceeded the mixing function, in contrast to only 2% of DIN
residuals. Residuals were shown relative to the DIP mixing function (y = − 0.06x + 2.2) which is zero on the y-axis. Additionally, the symbol colors reflect residual values relative to both
DIP and TDP (y = −0.09x + 3.3) mixing functions, under the assumption that all TDP from the MAR could be converted to DIP in the NGoM. DISCUSSION Over time, DIP concentrations significantly
increased in the NGoM, but with greater interannual variability over time. This rise in DIP was notable not only because of its increase over time, but also because DIP concentrations were
frequently in excess of the MAR DIP mixing function across the salinity gradient. Even when we used annual values of MAR DIP concentrations, about half of the NGoM DIP concentrations still
exceeded the mixing line. Thus, although MAR DIP loading rose over time, it was insufficient to explain the distribution of DIP concentrations in the NGoM. This contrasts to MAR DIN values,
which could account for NGoM DIN concentrations along the salinity gradient because DIN concentrations were predominantly (98%) below the MAR DIN mixing function. Other studies have
similarly proposed that there are excess sources of DIP (or selective removal of DIN) in the NGoM shelf region beyond the average MAR outflow area60,68. A two-endmember mixing model for NGoM
nutrient distributions is highly simplified and inputs from freshwater endmembers other than the MAR certainly contribute to NGoM nutrient concentrations69. Nevertheless, the high relative
contribution of the MAR to NGoM nutrient loading32 and the temporal correlations between MAR nutrient loads and NGoM concentrations shown in this study support our focus on the MAR as the
dominant freshwater endmember. Moreover, this two-endmember approximation does effectively highlight fundamental differences in the spatial and temporal patterns of DIN and DIP in the NGoM
that require further exploration. Resolving the NGoM DIP-salinity relationship requires a process that provides DIP to the NGoM in excess of the MAR DIP source along the entire salinity
gradient, that also increases through time, at least over the shelf region. In this context it is worth highlighting that the slopes of DIN, DIP, and DIN:DIP vs. salinity are nearly
identical across the hypoxic, shelf, and offshore regions, suggesting that the same processes control nutrient patterns regardless of distance from shore, or bottom depth. Within the MAR DIP
plume itself, relevant processes to consider include DIP regeneration and recycling, precipitation and aerosols, adsorption/desorption of P from suspended particles and benthic sediment,
vertical mixing, and DOP mineralization. DIP regeneration/recycling, the conversion of autochthonous organic P to DIP6, is an important source of P to NGoM phytoplankton, and P turnover
times within the upper water column are rapid70. However, such cycling cannot by itself explain NGoM DIP concentrations that surpass the MAR DIP mixing function since regeneration of DIP
from the MAR would need additional DIP sources to exceed mixing. Throughout the MAR watershed and NGoM, P is deposited from rainfall, dust, and anthropogenic emissions71,72. While
atmospheric P deposition has increased over time in other locations73, the atmospheric DIP and TP contribution to the NGoM is still very small (2–10 ng m−3)74. DIP adsorbs to particles in
freshwater, and is released with increasing salinity and discharge69,75. However, suspended-sediment-derived P is unlikely to explain the upward trend in NGoM DIP, because the percentage of
P in MAR suspended sediment stayed relatively stable over time (Fig. S4d), while MAR suspended sediment concentrations have significantly declined since 1985. P can also be remobilized into
bottom waters from benthic iron or sulfate reduction, mediated by oxygen levels76,77. For example, during NGoM hypoxic events, TDP is released into the water column from bottom sediments7.
Though benthic DIP sources are likely important within the MAR watershed and in shallow, nearshore areas of the NGoM69, our results show a pattern of excess DIP at high salinities, that are
typically found over deep water columns, further offshore. Although stratification in the NGoM can be strong78, the two-endmember model applied to the nutrient distributions implies
continuous vertical or horizontal mixing between the endmembers. While vertical mixing transports DIP to the surface from deeper depths, it also transports DIN. To provide excess DIP to the
NGoM system, the deeper waters being mixed upwards would need to have lower DIN:DIP than the offshore endmember. In addition, to align with the temporal trend in DIP, there would need to be
an increase in either the mixing intensity over time, or a decrease in DIN:DIP in those deeper waters. Further research will be needed to assess these possibilities. Biological
mineralization of organic P compounds through enzymatic reactions12,79 or photolysis80 could break down organic P-containing compounds originally delivered from the MAR, adding DIP to the
NGoM water column in excess of the MAR DIP mixing function. Figure 5 demonstrates that including MAR TDP inputs would be sufficient to explain most NGoM DIP concentrations if all MAR-derived
DOP was converted into DIP along the salinity gradient in the NGoM. Thus, the combination of DOP and DIP loading from the MAR could account for increasing DIP concentrations in the NGoM,
though other processes are not excluded. While it seems unlikely that all MAR-derived DOP would be converted to DIP, the match with the DIP data suggests that DOP mineralization is an
important source of DIP along the salinity gradient. Further research quantifying the fraction and processes of MAR DOP mineralization along the salinity gradient would help constrain the
degree to which MAR DOP inputs can explain NGoM DIP trends. This study expands upon previous nutrient research in the MAR and NGoM. From 1985 to 2019, N and P loading in the MAR increased
greatly relative to the 1950s33,36. During the period of this study, DIN loading fluctuated around a value of approximately 1 × 109 kg yr−1, while DON loading decreased significantly by
approximately 20% from 1985 to 2019. Stabilization of DIN loads60 and decreasing DON loads could reflect upstream N management (e.g., changing N fertilizer application timing)55. Conversely,
MAR DIP and DOP loading significantly increased from 1985 to 2019, contrasting with conclusions from studies based on earlier data40. Similar to many other NGoM nutrient studies, the NGoM
surface nutrient data set is dominated by samples collected in the summer months (Fig. S2). Increases in P in the MAR watershed could come from myriad sources over time, since the watershed
covers almost half the contiguous US states. Though P additions have been somewhat curtailed from wastewater treatment and industry, agriculture (i.e., pesticides, herbicides, and
fertilizers) is still a sizeable source of extremely high P inputs to the MAR81,82. Long-lived organic P species that are used in many pesticides, herbicides, and fertilizers have the
potential to persist in water and soil and act as a source of P that accumulates and is remobilized and recycled over long timescales83. In addition, legacy nutrients stored in soils and
reservoirs continually leach into the MAR watershed, and though legacy nutrient sources have been well studied for N13, the magnitude and residence times for legacy P pools are harder to
model and measure14,83. In the MAR, although there were clear temporal trends in DON loading and all forms of P loading, there was also considerable interannual variability. While MAR DIN
and DIP concentrations varied by approximately a factor of 2 interannually, MAR discharge ranged across nearly an order of magnitude over the same time period. MAR mean discharge in 2019 was
by far the highest of our study period, and also contributed the highest mean annual TDN and TDP loading since 1985. Prior studies have highlighted that much of the interannual variability
in MAR nutrient fluxes can be attributed to interannual variability in precipitation across the MAR watershed55,64,75. For example, 50 to 67% of the interannual variation in MAR N fluxes is
accounted for by river discharge alone60,84. In turn, MAR watershed precipitation and discharge has been shown to correlate with the El Niño Southern Oscillation and the Atlantic
Multidecadal Oscillation (AMO)34,56,60. In addition, during high flow years in which MAR spillways (e.g., Morganza, Bonnet Carré) were opened, nutrient dynamics and location of freshwater
delivery to the NGoM fundamentally change. The Bonnet Carré spillway diverts water from the Mississippi River into Lake Ponchartrain, which then connects to the NGoM, resulting in lower
concentrations of suspended sediment, TDN, TDP, DIN, and DIN:DIP, and higher DON content85. Openings of the Bonnet Carré spillway in 1997, 2008, 2011, 2016, 2018, and 201986,87, coincided
with low NGoM DIN:DIP in our study. However, it is difficult to assess the relative importance of spillway openings, versus general increased flow, on nutrient trends. Significant positive
temporal correlations between MAR nutrients, MAR loading, and MAR discharge, and NGoM nutrient concentrations and ratios indicated that processes within the MAR watershed, especially
influences on discharge, contributed significantly to interannual variability in NGoM surface nutrients, though with decreasing influence toward the offshore region. Similarly, Lehrter et
al. (2009) found that the influence of MAR discharge on chlorophyll concentration and primary productivity diminishes across the broader shelf. In the offshore region, interannual
variability in DIP concentrations remained significantly correlated to MAR discharge, MAR DIP concentrations, and MAR DIN:DIP, while interannual variability of offshore DIN concentrations
was not significantly correlated to the MAR observations. Thus, links between riverine inputs and offshore DIP concentrations were stronger than for offshore DIN, consistent with greater
nearshore N retention60,88. As mentioned above, the rise in MAR DIN loading and consequent nearshore hypoxia in the NGoM observed from the 1950s to the 1980s35,36,89 seems to have stabilized
during the period of this study. More recently, Karnauskas et al. (2015) suggested that an ecosystem-wide reorganization of the NGoM occurred in the mid-1990s as a result of physical
ecosystem changes driven by the AMO; which was in a cool phase from the 1970s to the 1990s, and in a warm phase from the mid-1990s to at least the mid-2000s. The AMO warm phase decreased
rainfall, MAR discharge, and mixed layer depth56. Some observations in this study may relate to the AMO because in the mid-1990s, annual mean NGoM DIN and DIP concentrations decreased from
earlier values, resulting in DIN:DIP close to and below 16; especially in the offshore region, where DIN and DIP concentrations were extremely low. In the early 2000s, DIN and DIP
concentrations began to rebound. If the AMO sets the stage for the processes that control nutrient distributions in the NGoM, as the AMO returns to a cool phase90 then we would expect DIP to
again fall. An extensive analysis of NGoM surface nutrient data from 1985 to 2012 by Cardona et al. (2016) suggested that there was an increase in surface DIN and DIP utilization from 2010
to 2011, possibly connected to the Deepwater Horizon oil spill, or to an exceptional deep winter mixed layer31. With the advantage of additional years of observations, the NGoM nutrient
conditions from 2010 to 2012 appear to have only been a temporary shift, as the prior conditions recurred post-2013. It remains to be seen whether the long-term increase in DIP
concentrations throughout the NGoM described in this study will lead to a longer-term shift in nutrients. Nutrient concentrations influence and correlate with phytoplankton biomass,
community composition, and spatial variance in chlorophyll-_a_ concentrations throughout the NGoM91,92. In this study, we showed that surface DIP concentrations in the NGoM increased over
time, while DIN did not. This increase in DIP relative to DIN has implications for nutrient limitation of phytoplankton growth and production, especially since the system has been classified
as both P and N limited through time11. While phytoplankton in the NGoM have been described as predominantly N limited11,31,52, transient P limitation (and higher DIN:DIP) has been observed
in spring and summer months in the MAR plume and nearshore region of the NGoM42,70,92. In general, N limitation typically occurs with N concentrations lower than 1 µM and N:P less than 10,
while P limitation commonly is associated with N:P greater than 3047,93,94, though empirical results sometimes deviate from the predictions of these criteria95. Nevertheless, applying these
nutrient ratio criteria, data in this study suggest P limitation is plausible throughout the salinity gradient in the NGoM, though the conditions for N limitation are more likely at high
salinities and in the offshore region45. Meanwhile, the temporal increase in NGoM DIP concentrations suggests more frequent and pervasive N limitation of phytoplankton over time. P
limitation at low salinities has the potential to lessen the effect of eutrophication on bottom water oxygen concentrations and reduce the incidence and strength of NGoM hypoxia54.
Increasing excess P may therefore increase the occurrence and severity of NGoM hypoxia. N fixation should be favored in N-limited surface waters with ample light and high DIP
concentrations96. Given increasing likelihood of N limitation and P availability, the suitability of the NGoM for N fixation may have increased during the 1985 to 2019 time period97. If this
trend continues, increased N fixation in the NGoM should also enhance C sequestration within the NGoM, as has been proposed for other areas affected by river plumes98. Nutrients in both the
MAR and the NGoM have changed over the last 35 years. In the MAR, N loading stabilized and P loading significantly increased since 1985. In the NGoM, DIP concentrations increased over time,
while DIN concentrations did not. These changes were primarily driven by trends in the shelf region, as opposed to further offshore. Increases in MAR DIP loading by itself could not explain
the distribution of NGoM DIP with salinity. MAR TDP loading would be sufficient to account for spatial and temporal DIP patterns in the NGoM, but only if all MAR DOP was converted to DIP by
enzymatic reactions and/or photolysis. Anomalous nutrient conditions from 2010 to 2012 did not persist. Nevertheless, the increasing likelihood of N limitation and excess P availability in
the NGoM has implications for phytoplankton communities, diazotrophy, and the prevalence and extent of nearshore hypoxia. DATA AVAILABILITY The datasets generated during and analyzed during
the current study are available from the corresponding author on reasonable request. All compiled data sources are included in the Supplementary Table S1. MAR nutrient concentration data
were compiled from USGS, and nutrient loads were sourced from USGS Load Estimation (LOADEST) and Weighted Regressions on Time, Discharge, and Season method (WRTDS) USGS data (Lee, 2022) from
St. Francisville, LA, USA (USGS 07373420 Hydrologic Unit 08070100). REFERENCES * Redfield, A. C. The biological control of chemical factors in the environment. _Am. Sci._ 46, 205–221
(1958). CAS Google Scholar * Howarth, R. Nutrient limitation of net primary production in marine ecosystems. _Annu. Rev. Ecol. Syst._ 19, 89–100 (1988). Article Google Scholar * Moore,
C. M. _et al._ Processes and patterns of oceanic nutrient limitation. _Nat. Geosci._ 6, 701–710 (2013). Article CAS ADS Google Scholar * Vitousek, P. M. & Howarth, R. W. Nitrogen
limitation on land and in the sea: How can it occur?. _Biogeochemistry_ 13, 87–115 (1991). Article Google Scholar * Tyrrell, T. The relative influences of nitrogen and phosphorus on
oceanic primary production. _Nature_ 400, 525–531 (1999). Article CAS ADS Google Scholar * Benitez-Nelson, C. R. The biogeochemical cycling of phosphorus in marine systems. _Earth Sci.
Rev._ 51, 109–135 (2000). Article CAS ADS Google Scholar * Adhikari, P. L., White, J. R., Maiti, K. & Nguyen, N. Phosphorus speciation and sedimentary phosphorus release from the
Gulf of Mexico sediments: Implication for hypoxia. _Estuar. Coast. Shelf Sci._ 164, 77–85 (2015). Article CAS Google Scholar * Geider, R. J. & La Roche, J. Redfield revisited:
variability of C:N: P in marine microalgae and its biochemical basis. _Eur. J. Phycol._ 37, 1–17 (2002). Article Google Scholar * Zehr, J. P. & Capone, D. G. Changing perspectives in
marine nitrogen fixation. _Science_ https://doi.org/10.1126/science.aay9514 (2020). Article PubMed Google Scholar * Carstensen, J. _et al._ Hypoxia in the Baltic Sea: Biogeochemical
cycles, benthic fauna, and management. _Ambio_ 43, 26–36 (2014). Article CAS PubMed PubMed Central ADS Google Scholar * Turner, R. E. & Rabalais, N. N. Nitrogen and phosphorus
phytoplankton growth limitation in the Northern Gulf of Mexico. _Aquat. Microb. Ecol._ 68, 159–169 (2013). Article Google Scholar * Duhamel, S. _et al._ Phosphorus as an integral component
of global marine biogeochemistry. _Nat. Geosci._ 14, 359–368 (2021). Article CAS ADS Google Scholar * Van Meter, K. J., Basu, N. B. & Van Capellen, P. Two centuries of nitrogen
dynamics: Legacy sources and sinks in the Mississippi and Susquehanna River Basins. _Glob. Biogeochem. Cycles_ 31, 2–23 (2017). Article Google Scholar * McCrackin, M. L. _et al._ A century
of legacy phosphorus dynamics in a large Drainage Basin. _Glob. Biogeochem. Cycles_ 32, 1107–1122 (2018). Article CAS ADS Google Scholar * Rabalais, N. N. _et al._ Nutrient-enhanced
productivity in the Northern Gulf of Mexico: Past, present and future. In _Geological Society Special Publication_ 39–63 (2002). https://doi.org/10.1007/978-94-017-2464-7_4. * Fennel, K.
& Laurent, A. N and P as ultimate and proximate limiting nutrients in the Northern Gulf of Mexico: Implications for hypoxia reduction strategies. _Biogeosciences_ 15, 3121–3131 (2018).
Article CAS ADS Google Scholar * Redfield, A.C. On the proportions of organic derivatives in sea water and their relation to the composition of plankton. James Johnstone Memorial Volume.
_Univ. Press Liverp._ 176–192 (1934). * Downing, J. A. Marine nitrogen: Phosphorus stoichiometry and the global N: P cycle. _Biogeochemistry_ 37, 237–252 (1997). Article Google Scholar *
Turner, R. E., Rabalais, N. N., Justic, D. & Dortch, Q. Global patterns of dissolved N, P and Si in large rivers. _Biogeochemistry_ 64, 297–317 (2003). Article CAS Google Scholar *
Wang, W. L., Moore, J. K., Martiny, A. C. & Primeau, F. W. Convergent estimates of marine nitrogen fixation. _Nature_ 566, 205–2011 (2019). Article CAS PubMed ADS Google Scholar *
Nausch, M. & Nausch, G. Phosphorus speciation and transformation along transects in the Benguela upwelling region. _J. Mar. Syst._ 140, 111–112 (2014). Article Google Scholar *
Martiny, A. C. _et al._ Biogeochemical controls of surface ocean phosphate. _Sci. Adv._ 5, 341–369 (2019). Article ADS Google Scholar * Cochrane, J. D. & Kelly, F. Low-frequency
circulation on the Texas-Louisiana continental shelf. _J. Geophys. Res._ 91, 10645–10655 (1986). Article ADS Google Scholar * Wiseman, W. J. & Garvine, R. W. Plumes and coastal
currents near large river mouths. _Estuaries_ 18, 509–517 (1995). Article Google Scholar * Dagg, M., Benner, R., Lohrenz, S. & Lawrence, D. Transformation of dissolved and particulate
materials on continental shelves influenced by large rivers: Plume processes. _Cont. Shelf Res._ 24, 833–858 (2004). Article ADS Google Scholar * Walker, N. D., Wiseman, W. J., Rouse, L.
J. & Babin, A. Effects of River discharge, wind stress, and slope eddies on circulation and the satellite-observed structure of the Mississippi River Plume. _J. Coast. Res._ 21,
1228–1244 (2005). Article Google Scholar * Schiller, R. V., Kourafalou, V. H., Hogan, P. & Walker, N. D. The dynamics of the Mississippi River plume: Impact of topography, wind and
offshore forcing on the fate of plume waters. _J. Geophys. Res. Oceans_ 116, 6029 (2011). Article ADS Google Scholar * Hitchcock, G. L. _et al._ Property fields in an effluent plume of
the Mississippi river. _J. Mar. Syst._ 12, 109–126 (1997). Article Google Scholar * Morey, S. L. _et al._ Export pathways for river discharged fresh water in the Northern Gulf of Mexico.
_J. Geophys. Res. Oceans_ 108, 3303 (2003). Article ADS Google Scholar * Rabalais, N. N., Turner, R. E., Wiseman, W. J. & Boesch, D. F. A brief summary of hypoxia on the Northern Gulf
of Mexico continental shelf: 1985–1988. _Geol. Soc. Spec. Publ._ 58, 35–47 (1991). Article ADS Google Scholar * Cardona, Y. _et al._ Highly variable nutrient concentrations in the
Northern Gulf of Mexico. _Deep-Sea Res. Part II Top. Stud. Oceanogr._ 129, 20–30 (2016). Article CAS ADS Google Scholar * Fry, B. _et al._ Carbon Dynamics on the louisiana continental
shelf and cross-shelf feeding of Hypoxia. _Estuaries Coasts_ 38, 703–721 (2015). Article CAS Google Scholar * Rabalais, N. N. & Turner, R. E. Oxygen depletion in the gulf of Mexico
adjacent to the Mississippi river. _Past Present Water Column Anoxia_ 70, 73–78 (2006). Google Scholar * Gomez, F. A. _et al._ ENSO-induced co-variability of Salinity, Plankton biomass and
coastal currents in the Northern Gulf of Mexico. _Sci. Rep._ 9, 1–10 (2019). Article ADS Google Scholar * Rabalais, N. N. & Turner, R. E. Gulf of Mexico Hypoxia: Past, present, and
future. _Limnol. Oceanogr. Bull._ 28, 117–124 (2019). Article Google Scholar * Turner, R. E. & Rabalais, N. N. Changes in Mississippi River water quality this century. _BioScience_ 41,
140–147 (1991). Article Google Scholar * David, M. B., Drinkwater, L. E. & McIsaac, G. F. Sources of nitrate yields in the Mississippi River Basin: Sources of nitrate yields in the
Mississippi River Basin. _J. Environ. Qual._ 39, 1657–1667 (2010). Article CAS PubMed Google Scholar * Rabalais, N. N. _et al._ Nutrient changes in the Mississippi River and system
responses on the adjacent continental shelf. _Estuaries_ 19, 386–407 (1996). Article CAS Google Scholar * Stets, E. G., Kelly, V. J. & Crawford, C. G. Regional and temporal
differences in nitrate trends discerned from long-term water quality monitoring data. _J. Am. Water Resour. Assoc._ 51, 1394–1407 (2015). Article ADS Google Scholar * Goolsby, D. A., _et
al._ Flux and sources of nutrients in the Mississippi-Atchafalaya River Basin: Topic 3 Report for the Integrated Assessment on Hypoxia in the Gulf of Mexico. Item Type monograph. In _Rep.
Integr. Assess. Hypoxia Gulf Mex. NOAA Coast. Ocean Program Decis. Anal. Ser._ 17, (1999). * Seitzinger, S. P. _et al._ Global river nutrient export: A scenario analysis of past and future
trends. _Glob. Biogeochem. Cycles_ https://doi.org/10.1029/2009GB003587 (2010). Article Google Scholar * Sylvan, J. B. _et al._ Phosphorus limits phytoplankton growth on the Louisiana
shelf during the period of hypoxia formation. _Environ. Sci. Technol._ 40, 7548–7553 (2006). Article CAS PubMed ADS Google Scholar * Sylvan, J. B., Quigg, A., Tozzi, S. & Ammerman,
J. W. Eutrophication-induced phosphorus limitation in the Mississippi River plume: Evidence from fast repetition rate fluorometry. _Limnol. Oceanogr._ 52, 2679–2685 (2007). Article CAS ADS
Google Scholar * Quigg, A. _et al._ Going west: Nutrient limitation of primary production in the Northern Gulf of Mexico and the importance of the Atchafalaya River. _Aquat. Geochem._ 17,
519–544 (2011). Article CAS Google Scholar * Lohrenz, S. E., Dagg, M. J. & Whitledge, T. E. Enhanced primary production at the plume/oceanic interface of the Mississippi River.
_Cont. Shelf Res._ 10, 639–664 (1990). Article ADS Google Scholar * Lohrenz, S. E., Fahnenstiel, G. L. & Redalje, D. G. Nutrients, irradiance, and mixing as factors regulating primary
production in coastal waters impacted by the Mississippi River plume. _Cont. Shelf Res._ 19(9), 1113–1141 (1999). Article ADS Google Scholar * Wysocki, L. A., Bianchi, T. S., Powell, R.
T. & Reuss, N. Spatial variability in the coupling of organic carbon, nutrients, and phytoplankton pigments in surface waters and sediments of the Mississippi River plume. _Estuar.
Coast. Shelf Sci._ 69, 47–63 (2006). Article CAS ADS Google Scholar * Rabalais, N. N. _et al._ Hypoxia in the Gulf of Mexico. _J. Environ. Qual._ 30, 320–329 (2001). Article CAS PubMed
Google Scholar * Eadie, B. J. _et al._ Records of nutrient-enhanced coastal ocean productivity in sediments from the Louisiana continental shelf. _Estuaries_ 17, 754–765 (1994). Article
CAS Google Scholar * Parsons, M. L. & Dortch, Q. Sedimentological evidence of an increase in Pseudo-nitzschia (Bacillariophyceae)abundance in response to coastal eutrophication.
_Limnol. Oceanogr._ 47, 551–558 (2002). Article ADS Google Scholar * Lehrter, J. C., Murrell, M. C. & Kurtz, J. C. Interactions between freshwater input, light, and phytoplankton
dynamics on the Louisiana continental shelf. _Cont. Shelf Res._ 29, 1861–1872 (2009). Article ADS Google Scholar * Turner, R. E., Rabalais, N. N., Alexander, R. B., Mcisaac, G. &
Howarth, R. W. Characterization of nutrient, organic carbon, and sediment loads and concentrations from the Mississippi River into the Northern Gulf of Mexico. _Estuaries Coasts_ 30, 773–790
(2007). Article CAS Google Scholar * Greene, R. M., Lehrter, J. C. & Hagy, J. D. Multiple regression models for hindcasting and forecasting midsummer hypoxia in the Gulf of Mexico.
_Ecol. Appl._ 19, 1161–1175 (2009). Article PubMed Google Scholar * Laurent, A. & Fennel, K. Simulated reduction of hypoxia in the Northern Gulf of Mexico due to phosphorus
limitation. _Elem. Sci. Anthr._ https://doi.org/10.12952/journal.elementa.000022 (2014). Article Google Scholar * Lu, C. _et al._ Increased extreme precipitation challenges nitrogen load
management to the Gulf of Mexico. _Nat. Commun. Earth Environ._ https://doi.org/10.1038/s43247-020-00020-7 (2020). Article Google Scholar * Karnauskas, M. _et al._ Evidence of
climate-driven ecosystem reorganization in the Gulf of Mexico. _Glob. Change Biol._ 21, 2554–2568 (2015). Article ADS Google Scholar * Pante, E. & Simon-Bouhet, B. marmap: A package
for importing, plotting and analyzing bathymetric and topographic data in R. _PLoS ONE_ 8, e73051 (2013). Article CAS PubMed PubMed Central ADS Google Scholar * Goolsby, D. A.,
Battaglin, W. A., Aulenbach, B. T. & Hooper, R. P. Nitrogen input to the Gulf of Mexico. _J. Environ. Qual._ 30, 329–336 (2001). Article CAS PubMed Google Scholar * Wawrik, B., Paul,
J. H., Bronk, D. A., John, D. & Gray, M. High rates of ammonium recycling drive phytoplankton productivity in the offshore Mississippi River plume. _Aquat. Microb. Ecol._ 35, 175
(2004). Article Google Scholar * Lohrenz, S. E., Redalje, D. G., Cai, W. J., Acker, J. & Dagg, M. A retrospective analysis of nutrients and phytoplankton productivity in the
Mississippi River plume. _Cont. Shelf Res._ 28, 1466–1475 (2008). Article ADS Google Scholar * Lee, C. J. Nutrient loads to the Gulf of Mexico produced by the USGS national water quality
network, 1968–2021. (2022). * U.S. Geological Survey. National Water Information System data (USGS Water Data for the Nation). (2022). * Goolsby, D. A., Battaglin, W. A., Aulenbach, B. T.
& Hooper, R. P. Nitrogen flux and sources in the Mississippi River Basin. _Sci. Total Environ._ 248, 75–86 (2000). Article CAS PubMed ADS Google Scholar * Donner, S. D. &
Scavia, D. How climate controls the flux of nitrogen by the Mississippi River and the development of hypoxia in the Gulf of Mexico. _Limnol. Oceanogr._ 52, 856–861 (2007). Article CAS ADS
Google Scholar * Lee, C. J., Murphy, J. C., Crawford, C. G. & Deacon, J. R. Methods for computing water-quality loads at sites in the U.S. Geological Survey National Water Quality
Network. (2017). * Cael, B. B., Bisson, K., Boss, E., Dutkiewicz, S. & Henson, S. Global climate-change trends detected in indicators of ocean ecology. _Nature_ 619, 551–554 (2023).
Article CAS PubMed PubMed Central ADS Google Scholar * Walker, N. D. Satellite assessment of Mississippi River plume variability: causes and predictability. _Remote Sens. Environ._ 58,
21–35 (1996). Article ADS Google Scholar * Bargu, S. _et al._ Mississippi River diversions and phytoplankton dynamics in deltaic Gulf of Mexico estuaries: A review. _Estuar. Coast. Shelf
Sci._ 221, 39–52 (2019). Article CAS ADS Google Scholar * He, S. & Xu, Y. J. Phosphorus fluxes from three coastalwatersheds under varied agriculture intensities to the Northern Gulf
of Mexico. _Water Switz._ 10, 816 (2018). Article Google Scholar * Popendorf, K. & Duhamel, S. Variable phosphorus uptake rates and allocation across microbial groups in the
oligotrophic Gulf of Mexico. _Environ. Microbiol._ 17, 3992–4006 (2015). Article CAS PubMed Google Scholar * Walsh, J. J. _et al._ Red tides in the Gulf of Mexico: Where, when, and why?.
_J. Geophys. Res._ https://doi.org/10.1029/2004JC002813 (2006). Article PubMed PubMed Central Google Scholar * Migon, C. & Sandroni, V. Phosphorus in rainwater: Partitioning inputs
and impact on the surface coastal ocean. _Limnol. Oceanogr._ 44, 1160–1165 (1999). Article CAS ADS Google Scholar * Pan, Y. _et al._ Enhanced atmospheric phosphorus deposition in Asia
and Europe in the past two decades. _Atmos. Ocean. Sci. Lett._ 14, 100051 (2021). Article Google Scholar * Mahowald, N. _et al._ Global distribution of atmospheric phosphorus sources,
concentrations and deposition rates, and anthropogenic impacts. _Glob. Biogeochem. Cycles_ https://doi.org/10.1029/2008GB003240 (2008). Article Google Scholar * Tan, Z. _et al._ Increased
extreme rains intensify erosional nitrogen and phosphorus fluxes to the Northern Gulf of Mexico in recent decades. _Environ. Res. Lett._ 16, 054080 (2021). Article CAS ADS Google Scholar
* Sutula, M., Bianchi, T. S. & Mckee, B. A. Effect of seasonal sediment storage in the lower Mississippi River on the flux of reactive particulate phosphorus to the Gulf of Mexico.
_Limnol. Oceanogr._ 49, 2223–2235 (2004). Article ADS Google Scholar * Ding, S. _et al._ In situ, high-resolution evidence for iron-coupled mobilization of phosphorus in sediments. _Sci.
Rep._ 6, 1–11 (2016). CAS Google Scholar * Obenour, D. R., Michalak, A. M., Zhou, Y. & Scavia, D. Quantifying the impacts of stratification and nutrient loading on hypoxia in the
Northern Gulf of Mexico. _Environ. Sci. Technol._ 46, 5489–5496 (2012). Article CAS PubMed PubMed Central ADS Google Scholar * Paytan, A. & McLaughlin, K. The oceanic phosphorus
cycle. _Chem. Rev._ 107, 563–576 (2007). Article CAS PubMed Google Scholar * Karl, D. M. & Björkman, K. Dynamics of dissolved organic phosphorus. In _Biogeochemistry of Marine
Dissolved Organic Matter_ 233–334 (Burlington Academic Press, 2015). * Hébert, M. P., Fugère, V. & Gonzalez, A. The overlooked impact of rising glyphosate use on phosphorus loading in
agricultural watersheds. _Front. Ecol. Environ._ 17, 48–56 (2019). Article Google Scholar * Turner, R. E., Milan, C. S., Swenson, E. M. & Lee, J. M. Peak chlorophyll a concentrations
in the lower Mississippi River from 1997 to 2018. _Limnol. Oceanogr._ 67, 703–712 (2022). Article CAS ADS Google Scholar * Sharpley, A. _et al._ Phosphorus legacy: Overcoming the effects
of past management practices to mitigate future water quality impairment. _J. Environ. Qual._ 42, 1308–1326 (2013). Article CAS PubMed Google Scholar * McIsaac, G. F., David, M. B.,
Gertner, G. Z. & Goolsby, D. A. Relating net nitrogen input in the Mississippi River Basin to nitrate flux in the lower Mississippi River. _J. Environ. Qual._ 31, 1610–1622 (2002).
Article CAS PubMed Google Scholar * Lane, R. R., Day, J. W., Kemp, G. P. & Demcheck, D. K. The 1994 experimental opening of the Bonnet Carre Spillway to divert Mississippi River
water into Lake Pontchartrain, Louisiana. _Ecol. Eng._ 17, 411–422 (2001). Article Google Scholar * Parra, S. M. _et al._ Bonnet Carré Spillway freshwater transport and corresponding
biochemical properties in the Mississippi Bight. _Cont. Shelf Res._ 199, 104114 (2020). Article Google Scholar * U.S. Army Corps of Engineers. Spillway Operation Information (2019). *
Howe, S., Miranda, C., Hayes, C. T., Letscher, R. T. & Knapp, A. N. The dual isotopic composition of nitrate in the gulf of Mexico and Florida straits. _J. Geophys. Res. Oceans_ 125,
e2020JC016047 (2020). Article CAS ADS Google Scholar * Turner, R. E., Rabalais, N. N. & Justic, D. Gulf of Mexico hypoxia: Alternate states and a legacy. _Environ. Sci. Technol._ 42,
2323–2327 (2008). Article CAS PubMed ADS Google Scholar * Frajka-Williams, E., Beaulieu, C. & Duchez, A. Emerging negative Atlantic Multidecadal Oscillation index in spite of warm
subtropics. _Sci. Rep._ 7, 11224 (2017). Article PubMed PubMed Central ADS Google Scholar * Chen, X., Lohrenz, S. E. & Wiesenburg, D. A. Distribution and controlling mechanisms of
primary production on the Louisiana-Texas continental shelf. _J. Mar. Syst._ 25, 179–207 (2000). Article Google Scholar * Qian, Y., Jochens, A. E., Kennicutt, M. C. & Biggs, D. C.
Spatial and temporal variability of phytoplankton biomass and community structure over the continental margin of the northeast Gulf of Mexico based on pigment analysis. _Cont. Shelf Res._
23, 1–17 (2003). Article ADS Google Scholar * Dortch, Q. & Whitledge, T. E. Does nitrogen or silicon limit phytoplankton production in the Mississippi River plume and nearby regions?.
_Cont. Shelf Res._ 12, 1293–1309 (1992). Article ADS Google Scholar * Goldman, J. C., McCarthy, J. J. & Peavey, D. G. Growth rate influence on the chemical composition of
phytoplankton in oceanic waters. _Nature_ 279, 210–215 (1979). Article CAS ADS Google Scholar * Juhl, A. & Murrell, M. Nutrient limitation of phytoplankton growth and physiology in a
subtropical estuary (Pensacola Bay, Florida). _Bull. Mar. Sci._ 82(1), 59–82 (2008). Google Scholar * Gruber, N. Elusive marine nitrogen fixation. _Proc. Natl. Acad. Sci. U.S.A._ 113,
4246–4248 (2016). Article CAS PubMed PubMed Central ADS Google Scholar * Mulholland, M. R., Berhardt, P. W., Heil, C. A., Bronk, D. A. & O’Neil, J. M. Nitrogen fixation and release
of fixed nitrogen by _Trichodesmium_ spp. in the Gulf of Mexico. _Limnol. Oceanogr._ 51, 1762–1776 (2006). Article CAS ADS Google Scholar * Subramaniam, A. _et al._ Amazon River
enhances diazotrophy and carbon sequestration in the tropical North Atlantic Ocean. _Proc. Natl. Acad. Sci. U.S.A._ 105, 10460–10465 (2008). Article CAS PubMed PubMed Central ADS Google
Scholar Download references ACKNOWLEDGEMENTS This study was supported by National Science Foundation award 1737240. We thank the captain and crew aboard R/V Endeavor on cruises EN620 and
EN642. We thank Benjamin Ramcharitar, Nicholas Bock, Ella Steiger, and John Richardson for their help with collection and analyses, Annalisa Bracco and Yuley Cardona for their gracious
sharing of data and guidance. We also thank Chief Scientist Joe Montoya, Anna Martin, and Erica Strope. We sincerely thank and are grateful to all the teams that collected and processed all
of the historical samples used in this study. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Lamont-Doherty Earth Observatory, Columbia University, 61 Route 9W, Palisades, NY, 10964, USA
Kailani G. Acosta, Andrew R. Juhl & Ajit Subramaniam * Department of Molecular and Cellular Biology, University of Arizona, Tucson, AZ, 85721, USA Solange Duhamel Authors * Kailani G.
Acosta View author publications You can also search for this author inPubMed Google Scholar * Andrew R. Juhl View author publications You can also search for this author inPubMed Google
Scholar * Ajit Subramaniam View author publications You can also search for this author inPubMed Google Scholar * Solange Duhamel View author publications You can also search for this author
inPubMed Google Scholar CONTRIBUTIONS K.A. contributed to conceptualization, methodology, formal analysis, investigation, data curation, writing original draft, writing, review, and
editing, and visualization. A.R.J. contributed to conceptualization, methodology, resources, writing original draft, writing, review, and editing, supervision, project administration, and
funding acquisition. A.S. and S.D. contributed to conceptualization, writing, review, and editing, and funding acquisition. All authors approved the submitted version. CORRESPONDING AUTHOR
Correspondence to Kailani G. Acosta. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains
neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This
article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as
you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party
material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the
article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Acosta, K.G., Juhl, A.R.,
Subramaniam, A. _et al._ Spatial and temporal variation in surface nitrate and phosphate in the Northern Gulf of Mexico over 35 years. _Sci Rep_ 14, 7305 (2024).
https://doi.org/10.1038/s41598-024-58044-4 Download citation * Received: 20 October 2023 * Accepted: 25 March 2024 * Published: 27 March 2024 * DOI:
https://doi.org/10.1038/s41598-024-58044-4 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not
currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative