Play all audios:
ABSTRACT Cardiac fibrosis contributes to the development of heart failure, and is the response of cardiac fibroblasts (CFs) to pressure or volume overload. Limiting factors in CFs research
are the poor availability of human cells and the tendency of CFs to transdifferentiate into myofibroblasts when cultured in vitro. The possibility to generate CFs from induced pluripotent
stem cells (iPSC), providing a nearly unlimited cell source, opens new possibilities. However, the behaviour of iPSC-CFs under mechanical stimulation has not been studied yet. Our study
aimed to assess the behaviour of iPSC-CFs under mechanical stretch and pro-fibrotic conditions. First, we confirm that iPSC-CFs are comparable to primary CFs at gene, protein and functional
level. Furthermore, iPSC-derived CFs adopt a pro-fibrotic response to transforming growth factor beta (TGF-β). In addition, mechanical stretch inhibits TGF-β-induced fibroblast activation in
iPSC-CFs. Thus, the responsiveness to cytokines and mechanical stimulation of iPSC-CFs demonstrates they possess key characteristics of primary CFs and may be useful for disease modelling.
SIMILAR CONTENT BEING VIEWED BY OTHERS CHRONIC ACTIVATION OF HUMAN CARDIAC FIBROBLASTS IN VITRO ATTENUATES THE REVERSIBILITY OF THE MYOFIBROBLAST PHENOTYPE Article Open access 26 July 2023
CARDIAC FIBROBLASTS AND MECHANOSENSATION IN HEART DEVELOPMENT, HEALTH AND DISEASE Article 14 November 2022 RECENT ADVANCES AND FUTURE PROSPECTS IN DIRECT CARDIAC REPROGRAMMING Article 11
December 2023 INTRODUCTION Heart failure is the most common cause of cardiovascular death, with a 5-year mortality above 50%1. One of the pathological changes resulting in heart failure is
cardiac fibrosis2. Cardiac fibrosis is one of the primary responses to acute injury (replacement fibrosis), such as cardiac infarction, and to chronic stress (interstitial fibrosis), such as
pressure overload. Pressure overload-induced cardiac fibrosis is commonly seen in systemic hypertension and pulmonary hypertension, in the left and right ventricles, respectively3,4. The
resulting interstitial cardiac fibrosis is characterized by accumulation and qualitative changes of the extracellular matrix (ECM). As a result, the passive stiffness of the ventricle
increases and its relaxation is impaired5,6,7. Even though cardiac fibrosis plays a significant role in heart failure, current therapies are unable to reverse cardiac fibrosis8. Cardiac
fibrosis involves the activation of cardiac fibroblasts (CFs)9. Under physiological conditions, CFs are responsible for the homeostasis of the ECM in the heart. The cardiac ECM is dynamic
and involves a constant balance between the production and degradation of ECM proteins. ECM proteins are produced as monomers and crosslinked to form strong fibers. Degradation of ECM
proteins is mediated by a balance between matrix metalloproteinases and their inhibitors. Under pathological conditions, such as pressure overload, CFs are further activated and will
transdifferentiate into cardiac myofibroblasts. Myofibroblasts are characterized by increased production and deposition of ECM proteins along with an upregulated expression of α-smooth
muscle actin (α-SMA). The transdifferentiation of CFs into myofibroblasts is heavily regulated by the transforming growth factor beta (TGF-β) pathway and mechanotransduction3. In fact,
stiffness of the cellular environment, shear stress and mechanical strain can be sensed through mechanosensitive complexes and affect the transdifferentiation10. While the importance of
mechanical stimuli has been acknowledged, studying the behaviour of CFs in vitro and developing anti-fibrotic treatments has remained challenging. The in vitro study of cardiac fibrosis is
limited by the poor availability of primary CFs and the fact that primary CFs quickly transdifferentiate into myofibroblasts when cultured on stiff, plastic culture plates and in the
presence of fetal bovine serum11,12. This limits the time cells isolated from tissue can be used to address scientific questions. In addition, when studying the development of heritable
heart diseases, healthy tissue to isolate fibroblasts from is often not available. When CFs are isolated from diseased tissue, human primary CFs are commonly in a pre-activated state and may
not represent the earlier stages of disease when fibrosis has not yet reached an irreversible end-stage. Therefore, instead of using primary CFs, induced pluripotent stem cells
(iPSC)-derived CFs may offer a suitable alternative. iPSCs are commonly used to generate various cell types, such as cardiomyocytes, to model diseases. Recently, several protocols have been
published to generate CFs13,14,15, providing the scientific community with a powerful tool to study the development of cardiac fibrosis. Although iPSC-CFs have been characterized and
compared to primary CFs, their behaviour under mechanically dynamic conditions has not been investigated yet. It has been shown that cyclic stretch of primary CFs at physiological levels can
alter the response to biochemical stimuli16,17,18. However, it is not known whether iPSC-CFs possess a similar kind of mechanosensitive response. To answer this question, this study aimed
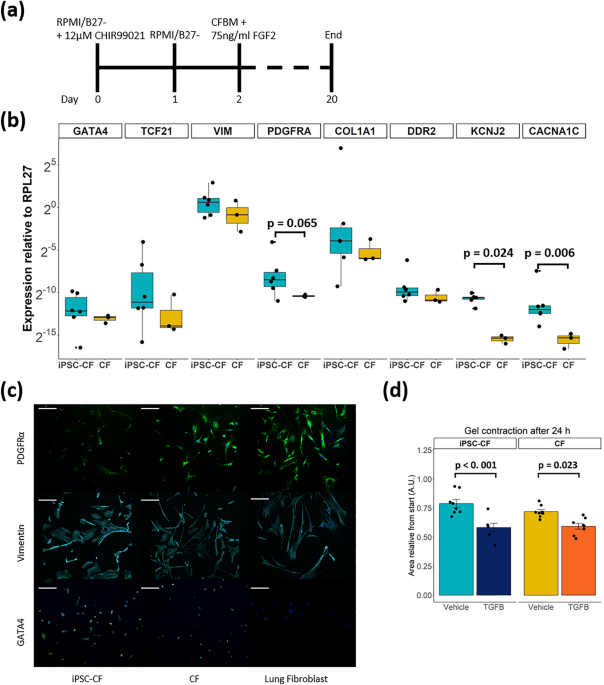
to study the effect of mechanical and pro-fibrotic stimulation on iPSC-derived CFs. RESULTS COMPARABLE GENE EXPRESSION OF CARDIAC FIBROBLAST MARKERS AND GEL CONTRACTION IN IPSC-CFS AND
PRIMARY CFS We generated iPSC-CFs following the protocol established by Zhang et al.13 (Fig. 1a). To ensure differentiation of iPSCs into iPSC-CFs the cells were characterized at the gene,
protein and functional level. At the end of differentiation, the cells expressed markers associated with the cardiac lineage, such as the genes encoding GATA4 (_GATA4_) and transcription
factor 21 (_TCF21_). In addition, fibroblast genes _VIM_, _PDGFRA_, _COL1A1_ and _DDR2_ were expressed at comparable levels as primary CFs (Fig. 1b). Gene expression of ion channels involved
in the conductance of the cardiac action potential (_KCNJ2, CACNA1C)_ was higher in iPSC-CFs compared to primary CFs. The morphology of the iPSC-CFs was spindle-shaped and comparable to
primary CFs. At the protein level, iPSC-CFs expressed the common fibroblast markers PDGFRα and vimentin, as do primary CFs and primary lung fibroblasts. Confirming their cardiac identity,
iPSC-CFs as well as primary CFs showed nuclear expression of the cardiac transcription factor GATA4 while this transcription factor was absent on primary lung fibroblasts (Fig. 1c). To
characterize the functionality of iPSC-CFs a gel contraction assay was performed, which is indicative for cell-ECM interaction. Similar to primary CFs, the iPSC-CFs contracted the gel which
was further increased when stimulating with TGF-β (Fig. 1d, Fig. S1). Overall, our data confirms that we generated functional CFs from human iPSCs which are comparable to primary human CFs.
STRETCH AND TGF-Β CO-STIMULATION REDUCED COLLAGEN 1 EXPRESSION IN IPSC-CFS To study how iPSC-CFs behave in a mechanically dynamic environment like the heart, cells were exposed to 10% cyclic
stretch at 1 Hz for 72 h (Fig. S2). This resembles the strain that cells experience under physiological conditions in a human heart at rest16,18,19. In addition, cells were treated with
TGF-β as a pro-fibrotic stimulus. Collagen 1 expression, the most abundant component of the cardiac ECM20, was investigated. Exposing iPSC-CFs to cyclic stretch did not alter _COL1A1_ gene
expression (Fig. 2a). Stimulation with TGF-β showed a clear trend of _COL1A1_ upregulation, while cyclic stretch in the presence of TGF-β significantly reduced the expression of _COL1A1_.
Immunofluorescence and western blot showed that stretch reduced collagen 1 protein expression in iPSC-CF, both in the presence and absence of TGF-β (Fig. 2b, c, Fig. S3). To summarize,
cyclic stretch inhibits the expression of collagen 1, especially in the presence of TGF-β. STRETCH REDUCES THE EXPRESSION OF ECM REMODELLING GENES IN IPSC-CFS One important function of
fibroblasts is to maintain ECM homeostasis by regulating the balance between ECM production, degradation and modification through expression of matrix metallopeptidases, their inhibitors and
lysyl oxidases, respectively21. Therefore, we investigated gene expression of key-players in ECM regulation in iPSC-CF under pro-fibrotic and cyclically stretched conditions. In iPSC-CFs,
cyclic stretch or TGF-β stimulation did not affect mRNA levels of the matrix metallopeptidase 1 gene (_MMP1_) and neither of its inhibitor, encoded by _TIMP1_ (Fig. 3a, b). In unstimulated
conditions, _LOX_ and _LOXL2_, genes associated with ECM crosslinking, were not affected by stretch, while during TGF-β stimulation cyclic stretched reduced _LOX_ and _LOXL2_ expression in
iPSC-CFs (Fig. 3c, d). CYCLIC STRETCH INHIBITS TGF-Β INDUCED TRANSDIFFERENTIATION OF IPSC-CF INTO MYOFIBROBLASTS Pathologic conditions such as pressure overload and the presence of
pro-fibrotic cytokines will activate CFs and transdifferentiate them into myofibroblasts3. This transdifferentiation is most commonly marked by an upregulation of α-smooth muscle actin
(α-SMA). To investigate cardiac fibroblast transdifferentiation, iPSC-CFs were exposed to cyclic stretch (Fig. S2) and/or stimulated with TGF-β. Cyclic stretch had no effect on _ACTA2_
expression, the gene encoding α-SMA (Fig. 4a). TGF-β stimulation promoted induction of _ACTA2 _expression, although high variation per iPSC-CF batch was observed. Co-stimulation of cyclic
stretch and TGF-β resulted in a significant reduction of _ACTA2_ expression compared to the static condition with TGF- β stimulation. iPSC-CFs present with very low basal levels of α-SMA
protein and no α-SMA stress fibres were observed under static and stretched conditions (Fig. 4b, c, Fig. S3). In line with the mRNA data, cyclic stretch alone had no effect on α-SMA protein
expression. When stimulated with TGF-β, α-SMA protein levels increased, and immunofluorescence imaging revealed stress fibre formation under static conditions. Stimulation with TGF-β in
combination with cyclic stretch significantly reduced α-SMA protein expression and stress fibres formation in iPSC-CFs. To conclude, TGF-β induced transdifferentiation of iPSC-CFs into
myofibroblasts can be inhibited by cyclic stretch. CYCLIC STRETCH INHIBITS TGF-Β INDUCED SIGNALLING IN IPSC-CFS TGF-β signalling is one of the most well-studied pathways involved in
fibroblast activation. In order to investigate whether this pathway is activated in response to mechanical stimulation, _PAI1_ and _TGFB1_ gene expression were analysed. The _PAI1_ gene is a
direct target of the transcription factors downstream TGF-β signalling, and commonly used as a marker for TGF-β pathway activation22. Under static conditions, TGF-β stimulation increased
_PAI1_ gene expression (not significant), while cyclic stretch abrogated TGF-β-induced _PAI1_ expression (Fig. 5a). TGF-β stimulation or cyclic stretch did not alter _TGFB1_ expression (Fig.
5b), indicating that the effect of mechanical stimulation on the TGF-β pathway is not regulated at the gene level, but may be regulated at the protein level instead. In conclusion, cyclic
stretch can inhibit TGF-β induced gene expression independent of _TGFB1_ expression. DISCUSSION CFs are the main contributors of cardiac fibrosis development3. The availability of human CFs
is limited hampering the field to move forward. To date, CFs can be generated from iPSCs, which could provide an unlimited source of human CFs13,14,15. However, the behaviour of iPSC-CFs in
relation to mechanical stimulation had not been investigated yet. In this study we demonstrated that iPSC-CFs are comparable to primary CFs with regard to the expression of key CF markers at
gene and protein levels. Expression of the cardiac markers _GATA4_ and _TCF21_ indicate the cardiac lineage of the cells. Furthermore, expression of the mesenchymal markers _VIM and PDGFRA_
as well as the ECM component _COL1A1_ and the collagen binding receptor _DDR2_ support their fibroblast phenotype. In addition, we showed that iPSC-CFs respond to pro-fibrotic and
mechanical stimulation. TGF-β induces CFs transdifferentiation into myofibroblasts and promotes ECM remodelling. Mechanical stimulation in the form of cyclic stretch at physiological levels
reduces collagen expression in iPSC-CFs. Interestingly, cyclic stretch also protects against TGF-β stimulation, preventing the cells from transdifferentiating into myofibroblasts. One can
only use iPSC-derived cells when they accurately represent their primary counterparts. Key characteristics of CFs are a defined mRNA profile, responsiveness to pro-fibrotic cytokines,
interaction with the ECM and mechanical sensitivity. iPSC-CFs generated using the protocol developed by Zhang et al. showed a comparable RNA sequencing profile in iPSC-CFs and primary CFs13.
Using our iPSCs lines, following the same protocol we generated iPSC-CFs with an mRNA profile comparable to primary CFs. Furthermore, at a functional level we demonstrated that iPSC-CFs
interact with their environment in a similar way as primary CFs, and respond to pro-fibrotic stimulation. These results indicate that iPSC-CFs possess several key characteristics of primary
CFs and may be suitable to investigate the behaviour of CFs and develop disease models of cardiac fibrosis. In order to investigate the behaviour of CFs in their native environment, we next
investigated the behaviour of iPSC-CFs under physiologically relevant conditions. In an effort to mimic the dynamic environment of the continuously beating heart, we investigated the effects
of cyclic mechanical stretch on iPSC-CFs. The importance of mechanical stimulation has been acknowledged, but the effects of mechanical stimulation on CFs remain controversial in in vitro
studies23. On one hand, it has been reported that cyclic stretch may induce transdifferentiation of CFs into myofibroblasts. On the other hand, it has been shown that cyclic stretch may have
a protective effect instead. One of the main factors influencing this controversy is the usage of cell sources from different species. As primary human CFs are limited in availability,
iPSC-CFs could provide a representative and stable source of cells to move forward. In order to study how iPSC-CFs and primary CFs behave in a mechanically dynamic environment similar to the
heart, cells were exposed to 10% cyclic stretch at 1 Hz for 72 h19. With this approach, we demonstrated that: Cyclic stretch alone inhibits expression of collagen 1 but does not affect
iPSC-CFs transdifferentiation or expression of matrix remodelling genes. In addition, cyclic stretch is protective against TGF-β mediated myofibroblast transdifferentiation in iPSC-CFs,
resulting in normalised expression of collagen 1, α-SMA and matrix remodelling genes such as _TIMP1_ and _MMP1_. The cause of the aforementioned controversy in literature regarding either
the pro-fibrotic or anti-fibrotic response of CFs to mechanical stimulation is hard to pin-point; experimental conditions vary widely between studies, such as cell origin, the duration of
the experiment, the surface coating and the presence of serum. A common trend in all those studies is that there may be a time-dependent response of stretch. It was shown in primary mouse
CFs that the response starts with an initial increase in phosphorylation of AKT, a downstream kinase involved in the transduction of mechanical stimuli24,25. At the gene level, it was shown
in primary rat CFs that there is an initial increase in fibrotic markers (i.e. _ACTA2, TGFB1, CTGF)_ after 4 h followed by a reduced increase after 24 h26. Roche et al. observed a similar
effect in primary rat CFs with an apparent reduced increase of _COL1A1_ gene expression after 48 h compared to 24 h27. 72 h of cyclic stretch was instead shown to inhibit TGF-β induced
fibroblast activation in primary human CFs16,18. Furthermore, it has been demonstrated that 96 h of cyclic stretch can promote or inhibit the response of primary mouse CFs to a broad
spectrum of biochemical stimuli, including TGF-β, angiotensin II, interleukin-1β and others17. Overall, it appears that longer stimulation results in a gradual decrease of an initial
pro-fibrotic response with eventually cells balancing the fibrotic response to the mechanically active environment in order to reach homeostasis. We may hypothesize that the duration of this
response curve is dependent on different factors, including the origin and age of the cells, their culture conditions (surface coating, substrate stiffness, or medium supplementation with
serum) and the presence of other cell types23. A clear association between mechanosensing and a response of CFs is apparent, but there is a need for a reproducible cell type to better
understand this phenomenon. TGF-β signalling is one of the main pathways involved in the activation of CFs and development of cardiac fibrosis28. Exposure of iPSC-CFs to TGF-β promotes the
expression of fibrotic and myofibroblast markers, such as α-SMA. When stretched however, this effect is diminished. How mechanical changes communicate with the TGF-β pathway is not well
understood. On one hand, mechanical strain has been shown in tissue to release active TGF-β from the ECM, which would promote fibroblast activation29. On the other hand, in this in vitro
study mechanical strain appears to inhibit fibroblast activation, indicating that there may be other mechanisms at play in this model. It is unknown whether this anti-fibrotic effect is
directly caused by interplay between mechanosensitive complexes and the TGF-β pathway. Mechanosensitive receptors such as integrins or mechanoresponsive factors such as YAP/TAZ may
communicate with the TGF-β pathway30,31. Alternatively, cyclic stretch may have an indirect effect, for example through internalization of extracellular receptors, altering the response to
ligand stimulation. Regardless, the field of mechanotransduction in CFs remains requires further investigation. While iPSCs have started a new era of research, the usage of these cells comes
with limitations. iPSC-CFs showed many similarities with primary CFs, but the maturity of iPSC-derived cell lineages remains an important topic of contention. Although maturation is clearly
defined for some cell types, such as cardiac myocytes, a clear definition lacks for CFs. The heterogeneity and plasticity of this cellular population under physiological conditions makes it
difficult to set well defined standards of “mature” CFs32. iPSC-CFs present with various characteristics of primary cells, but they differ in several aspects as well. For example, Zhang et
al. noted an increased proliferation capacity in iPSC-CFs and foetal CFs compared to adult CFs, indicating the iPSC-CFs may be more foetal-like13. This increased proliferation capacity and
ability to stay in an inactivated state while in culture increases the applicability of the iPSC-CFs in research, as it has been demonstrated that CFs which have transdifferentiated into
myofibroblast will have an altered response to mechanical stimulation33. In addition, little is known about the electrophysiological characteristics of iPSC-CFs and their interaction with
other conducting cells such as cardiomyocytes34. Further electrophysiological characterisation should be performed to better understand the behaviour of these cell in the electrical circuit
of the heart. To conclude, in this study we demonstrated that iPSC-derived CFs show similar gene and protein expression as primary CFs. In addition, pro-fibrotic stimulation promoted
transdifferentiation of iPSC-CFs into a myofibroblast phenotype. When stimulated with cyclic stretch, this transdifferentiation is inhibited. Together, the mechano- and TGF-β-responsive
characteristics support the use of iPSC-CFs for physiological relevant disease modelling. Future studies could further dive into the mechanisms driving cardiac fibroblast behaviour and
cardiac fibrosis. METHODS CELL CULTURE Three human iPSCs lines were derived from one healthy female subject generated as described before35. iPSCs were cultured on vitronectin XF (StemCell
Technologies) coated plates in TeSR-E8 medium (StemCell Technologies). Cells were passaged every 7 days in a 1:10 ratio using 0.5 µM EDTA solution (Invitrogen) at RT for 5 min and manual
dissociation. Commercially available human primary ventricular cardiac fibroblasts (Lonza) from 1 healthy donor were cultured according to the manufacturer’s instructions. Primary cardiac
fibroblasts were used for a maximum of 3 passages. Mycoplasma tests were routinely performed for all cell cultures in this study. CARDIAC FIBROBLAST DIFFERENTIATION For the generation of
iPSC-CFs a protocol developed by Zhang et al. was used13. Briefly, human iPSCs were dissociated with 1 mL/well 0.5 µM EDTA solution (Invitrogen) at RT for 5 min and seeded on Vitronectin XF
(StemCell Technologies) coated 6-well plates at a density of 15.000–30.000 cells/cm2 in TeSR-E8 medium (StemCell Technologies) supplemented with 5 μM ROCK inhibitor (Y-27632) (Tocris) for 24
h. Cells were cultured for 6–7 days in TeSR-E8 medium with medium changes every other day until they reached 100% confluency and differentiation started (day 0). At day 0, the medium was
changed to 2.5 mL/well RPMI + B27 without insulin (Gibco) and supplemented with 12 µM CHIR99021 (Tocris) for 24 h (day 1). After day 1, the medium was changed to 2.5 mL RPMI + B27 without
insulin for 24 h (day 2). Afterwards, the medium was changed to 2.5 mL/well of the CFBM medium (Table S1) supplemented with 75 ng/mL bFGF (StemCell Technologies). Cells were refreshed with 2
mL/well CFBM supplemented with 75 ng/mL bFGF every other day until day 20 when RNA was collected, and cells were dissociated using TrypLE Select (10x) (Thermo Fisher) for 10 min at 37 °C.
After dissociation, cells were cultured in DMEM + 10% Fetal bovine serum. For the first two passages, 5 μM ROCK inhibitor was added for 24 h to help cell attachment. Cells between passage
3–6 were used for experiments. IMMUNOFLUORESCENT STAINING Cells were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 20 min at room temperature. Samples were
permeabilized, blocked, and incubated at 4 °C overnight with primary antibodies against GATA4 (1:200, Abcam, ab124265), PDGFR-α (1:250, Cell Signaling Technology, #3174), vimentin (1:500,
Abcam, ab73159) or α-SMA–Cy3-conjugated (1:400, Sigma, C6198). Samples were then incubated for 1 h at room temperature with secondary antibodies conjugated with Alexa-488, Alexa-555, or
Alexa-647 (1:500; Abcam). The cells were counterstained with Hoechst 33342 nuclear dye (1:500; Santa Cruz Biotechnology) and actiStain phalloidin conjugated to Alexa-670 (1:200; Cytoskeleton
Inc.). Images were captured under a laser confocal microscope (Nikon A1R, Nikon; RCM1, confocal.nl) and analysed on ImageJ (NIH). QUANTITATIVE REAL-TIME PCR ANALYSIS Total RNA was extracted
using the RNeasy® Mini Kit with DNAse I digestion (Qiagen) following the manufacturer’s protocol. The concentration and purity of the RNA were measured on the Nanodrop One spectrophotometer
(Thermofisher Scientific). Reverse transcription to cDNA was carried out using the iScript™ cDNA Synthesis Kit (Bio-Rad). Quantitative real-time PCR amplifications were performed with 10 ng
(2 µL) of cDNA in a final volume of 10 µL, containing 5 µL of Fast SYBR Green MasterMix (Thermofisher Scientific), 2 µL of RNase-free PCR-grade water (Thermofisher Scientific), and 1 µL of
both forward and reverse 10 μM primer solutions (sequences specified in Table S2). Housekeeping gene ribosomal protein L27 (RPL27) was used to ensure the validity and reproducibility of the
results. Data were collected and analyzed in duplicate on the CFX384™ Real-Time System (C1000 Touch™ Thermal Cycler, Bio-Rad). The Livak method was used to quantify the relative (2−ΔΔCT)
expression of each gene between groups. WESTERN BLOT Whole cell lysate samples were collected in lysis buffer (20 mM Tris–HCl, 150 mM NaCl, 100 mM KCl, 2 mM EDTA-NaOH, 5% Igepal, and 0.5%
Triton X-100; pH 8.0) supplemented with PhosSTOP™ (Roche) and cOmplete™ Protease Inhibitor Cocktail (Roche) according to manufacturer’s instructions. Lysates were prepared with 1 × NuPage
LDS sample buffer (Thermofisher Scientific) and 50 µM DTT (Thermofisher Scientific). Protein samples were loaded on 4–12% NuPage Bis–Tris protein gel (Thermofisher Scientific) and
electrophoresed at 200 V for ~ 1 h. Separated proteins were transferred to 0.45 µM Amersham Hybond ECL nitrocellulose membranes (Thermofisher Scientific) and blocked with 5% BSA (Sigma) in
Tris-buffered saline (pH 7.6) with 0.1% Tween (TBS-T) for 1 h at room temperature. Membranes were incubated overnight at 4 °C with gentle shaking in either 5% BSA or 5% milk with primary
antibodies against collagen 1 (1:2000, Abcam, Ab138492), vinculin (1:1000, Sigma-Aldrich, V9131), α-SMA (1:1000, Dako, M0851). Afterwards, appropriate HRP-conjugated secondary antibodies
were incubated in 5% BSA or 5% milk for 1 h at room temperature. Bands were visualized with Amersham ECL Prime Blotting Detection Reagent (GE Healthcare Life Sciences) detected with the
Amersham™ Imager 600 (GE Healthcare Life Sciences), quantified with ImageJ (NIH), and normalized to vinculin expressions. GEL CONTRACTION ASSAY Cells were mixed with rat tail collagen type 1
(Corning) according to the manufacturer’s instructions to a final concentration of 2 mg/mL, in a final volume of 150 μl and 50.000 cells per gel. The mixture was pipetted onto a glass
coverslip in a 6-well plate and allowed to harden for 30 min at 37 °C. Next, 1.5 mL DMEM + 10% fetal bovine serum was added to the well to cover the gel. The following day, the medium was
replaced with DMEM and the cells were starved for 6 h. Afterwards, the gels were detached from the coverslips and medium was replaced with either DMEM or DMEM + 10 ng/mL TGF-β1 (Stem Cell
Technologies). Pictures were taken after 24 h to monitor the contraction of the gels. MECHANICAL STRETCH AND TGF-Β STIMULATION Cells were seeded on rat tail collagen I coated BioFlex®
Culture Plates (Dunnlab) coated with 5 μg/ml fibronectin (Sigma) according to the manufacturer’s instructions, using DMEM with 10% fetal bovine serum. After 24 h, the medium was changed
either to DMEM or DMEM + 10 ng/mL TGF-β1 and cells were subjected to equiaxial cyclic strain in a sinusoidal pattern (0–10%) at 1 Hz for 72 h using a Flexcell FX-6000 Tension straining
device (Flexcell, Dunnlab) and compared to static cultures (0%). Strain experiments were performed with 6 different batches of CFs differentiations, using 3 iPSCs clones of one healthy
subject. STATISTICAL ANALYSIS Statistical analyses were performed with R statistics package version 4.2.3 (R Foundation for Statistical Computing, Vienna, Austria) and R studio version
2023.09.0 Build 463 (R-studio, Boston, MA, USA). Normality of the data was checked using a Shapiro–Wilk test with _p_-values < 0.05 considered as not normal. Data from _KCNJ2_ gene
expression and collagen 1 protein expression was not normally distributed. Unless otherwise indicates statistics were performed as follows. When comparing two groups, a student’s t-test was
applied. In the case of _KCNJ2_, the non-parametric Mann–Whitney U test was applied. When comparing multiple groups, a linear mixed model, with the experimental conditions considered as
fixed parameters and the batches of differentiation as random parameters was applied. Tukey post-hoc tests were performed to correct for multiple comparisons. For collagen 1 protein
expression the non-parametric Kruskal–Wallis test was performed. _P_-values < 0.05 were considered statistically significant. DATA AVAILABILITY No datasets were generated or analysed
during the current study. CHANGE HISTORY * _ 10 JUNE 2024 A Correction to this paper has been published: https://doi.org/10.1038/s41598-024-64127-z _ REFERENCES * Tsao, C. W. _et al._ Heart
disease and stroke statistics—2022 update: A report from the american heart association. _Circulation_ 145, e153–e639 (2022). Article PubMed Google Scholar * González, A., Schelbert, E.
B., Díez, J. & Butler, J. Myocardial interstitial fibrosis in heart failure: Biological and translational perspectives. _J. Am. Coll. Cardiol._ 71, 1696–1706 (2018). Article PubMed
Google Scholar * Frangogiannis, N. G. Cardiac fibrosis. _Cardiovasc. Res._ 117, 1450–1488 (2021). Article CAS PubMed Google Scholar * Bekedam, F. T., Goumans, M. J., Bogaard, H. J., de
Man, F. S. & Llucià-Valldeperas, A. Molecular mechanisms and targets of right ventricular fibrosis in pulmonary hypertension. _Pharmacol. Ther._ 244, 108389 (2023). Article CAS PubMed
Google Scholar * Chaturvedi, R. R. _et al._ Passive stiffness of myocardium from congenital heart disease and implications for diastole. _Circulation_ 121, 979–988 (2010). Article PubMed
Google Scholar * Yamamoto, K. _et al._ Myocardial stiffness is determined by ventricular fibrosis, but not by compensatory or excessive hypertrophy in hypertensive heart. _Cardiovasc.
Res._ 55, 76–82 (2002). Article CAS PubMed Google Scholar * Rain, S. _et al._ Right ventricular diastolic impairment in patients with pulmonary arterial hypertension. _Circulation_ 128,
2016–2025 (2013). Article CAS PubMed Google Scholar * de Boer, R. A. _et al._ Towards better definition, quantification and treatment of fibrosis in heart failure. A scientific roadmap
by the committee of translational research of the heart failure association (HFA) of the European society of cardiology. _Eur. J. Heart Fail._ 21, 272–285 (2019). Article PubMed Google
Scholar * Travers, J. G., Kamal, F. A., Robbins, J., Yutzey, K. E. & Blaxall, B. C. Cardiac Fibrosis: The fibroblast awakens. _Circ. Res._ 118, 1021–1040 (2016). Article CAS PubMed
PubMed Central Google Scholar * Herum, K. M., Lunde, I. G., McCulloch, A. D. & Christensen, G. The soft- and hard-heartedness of cardiac fibroblasts: Mechanotransduction signaling
pathways in fibrosis of the heart. _J. Clin. Med._ 6, 53 (2017). Article PubMed PubMed Central Google Scholar * van Putten, S., Shafieyan, Y. & Hinz, B. Mechanical control of cardiac
myofibroblasts. _J. Mol. Cell. Cardiol._ 93, 133–142 (2016). Article PubMed Google Scholar * Baranyi, U. _et al._ Primary human fibroblasts in culture switch to a myofibroblast-like
phenotype independently of TGF beta. _Cells_ 8, 721 (2019). Article CAS PubMed PubMed Central Google Scholar * Zhang, J. _et al._ Functional cardiac fibroblasts derived from human
pluripotent stem cells via second heart field progenitors. _Nat. Commun._ 10, 2238 (2019). Article ADS PubMed PubMed Central Google Scholar * Zhang, H. _et al._ Generation of quiescent
cardiac fibroblasts from human induced pluripotent stem cells for in vitro modeling of cardiac fibrosis. _Circ. Res._ 125, 552–566 (2019). Article ADS CAS PubMed PubMed Central Google
Scholar * Whitehead, A. J., Hocker, J. D., Ren, B. & Engler, A. J. Improved epicardial cardiac fibroblast generation from iPSCs. _J. Mol. Cell. Cardiol._ 164, 58–68 (2022). Article CAS
PubMed Google Scholar * Watson, C. J. _et al._ Mechanical stretch up-regulates the B-type natriuretic peptide system in human cardiac fibroblasts: A possible defense against transforming
growth factor-β mediated fibrosis. _Fibrogenesis Tissue Repair_ 5, 9 (2012). Article CAS PubMed PubMed Central Google Scholar * Rogers, J. D., Holmes, J. W., Saucerman, J. J. &
Richardson, W. J. Mechano-chemo signaling interactions modulate matrix production by cardiac fibroblasts. _Matrix Biol. Plus_ 10, 100055 (2021). Article CAS PubMed Google Scholar *
Bracco Gartner, T. C. L. _et al._ Cyclic strain has antifibrotic effects on the human cardiac fibroblast transcriptome in a human cardiac fibrosis-on-a-chip platform. _J. Mech. Behav.
Biomed. Mater._ 144, 105980 (2023). Article CAS PubMed Google Scholar * Gupta, V. & Grande-Allen, K. J. Effects of static and cyclic loading in regulating extracellular matrix
synthesis by cardiovascular cells. _Cardiovasc. Res._ 72, 375–383 (2006). Article CAS PubMed Google Scholar * Bashey, R. I., Martinez-Hernandez, A. & Jimenez, S. A. Isolation,
characterization, and localization of cardiac collagen type VI. Associations with other extracellular matrix components. _Circ. Res._ 70, 1006–1017 (1992). Article CAS PubMed Google
Scholar * Rienks, M., Papageorgiou, A.-P., Frangogiannis, N. G. & Heymans, S. Myocardial extracellular matrix: An ever-changing and diverse entity. _Circ. Res._ 114, 872–888 (2014).
Article CAS PubMed Google Scholar * Keeton, M. R., Curriden, S. A., van Zonneveld, A. J. & Loskutoff, D. J. Identification of regulatory sequences in the type 1 plasminogen activator
inhibitor gene responsive to transforming growth factor beta. _J. Biol. Chem._ 266, 23048–23052 (1991). Article CAS PubMed Google Scholar * Bracco Gartner, T. C. L. _et al._ Advanced in
vitro modeling to study the paradox of mechanically induced cardiac fibrosis. _Tissue Eng. Part C. Methods_ 27, 100–114 (2021). Article PubMed Google Scholar * Suzuki, K. _et al._
Basigin promotes cardiac fibrosis and failure in response to chronic pressure overload in mice. _Arterioscler. Thromb. Vasc. Biol._ 36, 636–646 (2016). Article CAS PubMed Google Scholar
* Schmidt, J. B., Chen, K. & Tranquillo, R. T. Effects of intermittent and incremental cyclic stretch on ERK signaling and collagen production in engineered tissue. _Cell. Mol. Bioeng._
9, 55–64 (2016). Article CAS PubMed Google Scholar * Ploeg, M. C. _et al._ Piezo1 mechanosensitive ion channel mediates stretch-induced Nppb expression in adult rat cardiac fibroblasts.
_Cells_ 10, 1745 (2021). Article CAS PubMed PubMed Central Google Scholar * Roche, P. L. _et al._ Role of scleraxis in mechanical stretch-mediated regulation of cardiac myofibroblast
phenotype. _Am. J. Physiol. Cell Physiol._ 311, 297–307 (2016). Article Google Scholar * Yousefi, F. _et al._ TGF-β and WNT signaling pathways in cardiac fibrosis: non-coding RNAs come
into focus. _Cell Commun. Signal._ 18, 87 (2020). Article CAS PubMed PubMed Central Google Scholar * Hinz, B. The extracellular matrix and transforming growth factor-β1: Tale of a
strained relationship. _Matrix Biol._ 47, 54–65 (2015). Article CAS PubMed Google Scholar * Munger, J. S. & Sheppard, D. Cross talk among TGF-β signaling pathways, integrins, and the
extracellular matrix. _Cold Spring Harb. Perspect. Biol._ 3, a005017 (2011). Article PubMed PubMed Central Google Scholar * Ragazzini, S. _et al._ Mechanosensor YAP cooperates with
TGF-β1 signaling to promote myofibroblast activation and matrix stiffening in a 3D model of human cardiac fibrosis. _Acta Biomater._ 152, 300–312 (2022). Article CAS PubMed Google Scholar
* Wang, L. _et al._ Single-cell dual-omics reveals the transcriptomic and epigenomic diversity of cardiac non-myocytes. _Cardiovasc. Res._ 118, 1548–1563 (2022). Article CAS PubMed
Google Scholar * Kong, M. _et al._ Cardiac fibrotic remodeling on a chip with dynamic mechanical stimulation. _Adv. Healthc. Mater._ 8, e1801146 (2019). Article PubMed PubMed Central
Google Scholar * Hall, C., Gehmlich, K., Denning, C. & Pavlovic, D. Complex relationship between cardiac fibroblasts and cardiomyocytes in health and disease. _J. Am. Heart Assoc._ 10,
e019338 (2021). Article CAS PubMed PubMed Central Google Scholar * Llucià-Valldeperas, A. _et al._ Development of a 3-dimensional model to study right heart dysfunction in pulmonary
arterial hypertension: First observations. _Cells_ 10, 3595 (2021). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS Drs Bogaard, Vonk Noordegraaf,
Goumans, and de Man were supported by the Netherlands CardioVascular Research Initiative: the Dutch Heart Foundation, Dutch Federation of University Medical Centres, the Netherlands
Organisation for Health Research and Development, and the Royal Netherlands Academy of Sciences (CVON-2012-08 PHAEDRA, CVON-2018-29 PHAEDRA-IMPACT, and CVON-2017-10 Dolphin-Genesis). Drs
Aman, Vonk Noordegraaf and de Man were further supported by the Netherlands Organization for Scientific Research (NWO-VENI: 0915.01649.10155, NWO-VICI: 918.16.610, NWO-VIDI: 917.18.338). Dr
de Man was supported by a Dutch Heart Foundation Dekker senior postdoc grant (2018T059). Dr Aman was supported by the Dutch Lung Fund. Drs Bogaard and Vonk Noordegraaf have received research
grant support from Actelion, GSK, and Ferrer (Therabel). Dr de Man has received research grant support from Janssen and BIAL. All other authors have reported that they have no relationships
relevant to the contents of this paper to disclose. The authors thank the Microscopy & Cytometry Core Facility of the Amsterdam UMC for their help and usage of equipment. Schematic
figure was created using BioRender. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * PHEniX Laboratory, Department of Pulmonary Medicine, Amsterdam UMC Location Vrije Universiteit Amsterdam, De
Boelelaan 1117, Amsterdam, The Netherlands Fjodor T. Bekedam, Rowan Smal, Marisa C. Smit, Jurjan Aman, Anton Vonk-Noordegraaf, Harm Jan Bogaard, Frances S. De Man & Aida
Llucià-Valldeperas * Amsterdam Cardiovascular Sciences, Pulmonary Hypertension and Thrombosis, Amsterdam, The Netherlands Fjodor T. Bekedam, Rowan Smal, Marisa C. Smit, Jurjan Aman, Anton
Vonk-Noordegraaf, Harm Jan Bogaard, Frances S. De Man & Aida Llucià-Valldeperas * Department of Cell and Chemical Biology, Leiden UMC, 2300 RC, Leiden, The Netherlands Marie José Goumans
Authors * Fjodor T. Bekedam View author publications You can also search for this author inPubMed Google Scholar * Rowan Smal View author publications You can also search for this author
inPubMed Google Scholar * Marisa C. Smit View author publications You can also search for this author inPubMed Google Scholar * Jurjan Aman View author publications You can also search for
this author inPubMed Google Scholar * Anton Vonk-Noordegraaf View author publications You can also search for this author inPubMed Google Scholar * Harm Jan Bogaard View author publications
You can also search for this author inPubMed Google Scholar * Marie José Goumans View author publications You can also search for this author inPubMed Google Scholar * Frances S. De Man View
author publications You can also search for this author inPubMed Google Scholar * Aida Llucià-Valldeperas View author publications You can also search for this author inPubMed Google
Scholar CONTRIBUTIONS F. T. B.: writing manuscript, performing experiments & data analysis. R. S.: performing experiments. M. C. S.: performing experiments. J. A.: revised manuscript. A.
V. N.: revising manuscript & acquired funding. H. J. B: Revising manuscript & acquired funding. M. J. G.: supervision, revising manuscript & acquired funding. F. S. D. M.:
supervision, revising manuscript, & acquired funding. A. L. V.: supervision, revising manuscript. CORRESPONDING AUTHORS Correspondence to Frances S. De Man or Aida Llucià-Valldeperas.
ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional
claims in published maps and institutional affiliations. The original online version of this Article was revised: The original version of this Article contained an error in the title of the
paper, where the term “cell” was erroneously omitted. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative
Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the
original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in
the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your
intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence,
visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Bekedam, F.T., Smal, R., Smit, M.C. _et al._ Mechanical stimulation of
induced pluripotent stem cell derived cardiac fibroblasts. _Sci Rep_ 14, 9795 (2024). https://doi.org/10.1038/s41598-024-60102-w Download citation * Received: 15 December 2023 * Accepted: 18
April 2024 * Published: 29 April 2024 * DOI: https://doi.org/10.1038/s41598-024-60102-w SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get
shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative