Play all audios:
ABSTRACT Tartrazine (E102) is a controversial coloring agent whose potential impacts on human health are not fully understood. Our study reveals the vascular disrupting effects of tartrazine
(TTZ) on developing zebrafish embryos in vivo and on human umbilical vein endothelial cells in vitro. The dye was shown to cause dose-dependent hemorrhages in zebrafish embryos. Analyzing
transgenic zebrafish harboring fluorescent endothelial cells revealed that TTZ treatment disrupted cell organization into vessels in both the sub-intestinal vein and the brain area. Assays
on human umbilical vein endothelial cells demonstrated that TTZ inhibited endothelial proliferation, tube formation, and migration in a dose-dependent manner. Taken together, our results
indicate for the first time that TTZ can affect endothelial cell properties, possibly by disrupting Rho family GTPase pathways which control the cytoskeleton. Our finding provides a credible
explanation for many reported human health impacts and offers prospective applications for biomedicine. SIMILAR CONTENT BEING VIEWED BY OTHERS DEVELOPMENTAL TOXICITY AND MECHANISM OF
DIBUTYL PHTHALATE ON THE DEVELOPMENT OF SUBINTESTINAL VESSELS IN ZEBRAFISH Article Open access 18 November 2024 EXPOSURE TO THE NATURAL ALKALOID BERBERINE AFFECTS CARDIOVASCULAR SYSTEM
MORPHOGENESIS AND FUNCTIONALITY DURING ZEBRAFISH DEVELOPMENT Article Open access 15 October 2020 WULIANGYE _BAIJIU_ BUT NOT ETHANOL REDUCES CARDIOVASCULAR DISEASE RISKS IN A ZEBRAFISH
THROMBOSIS MODEL Article Open access 05 December 2022 INTRODUCTION Tartrazine (E102, TTZ), a lemon-yellow monoazo dye, is one of the most widely used coloring agents in food products,
medicines, and cosmetics. Despite being extensively studied in recent decades, TTZ remains among the most controversial food additives, with various debates over its potential safety and
toxicity1,2,3. The 2009 EFSA Panel on food additives set its acceptable daily intake (ADI) to 7.5 mg/kg bw based on the “no observed adverse effect level” (NOAEL) at 750 mg/kg bw/day
determined in a rat chronic toxicity assay1, while the Joint FAO/WHO expert committee in 2017 approved a higher ADI at 10 mg/kg bw3. However, recent studies have challenged the safety of TTZ
even at those ADI levels or lower. For instance, 30-day TTZ feeding to rats at 7.5 mg/kg bw induced liver and kidney histopathology, in addition to leucocyte DNA damage4. Another study
described fetal mortality and teratogenicity following 0.45 and 4.5 mg/kg bw TTZ administration to pregnant rats on the 6th −15th days of gestation5. Similarly, neurotoxic6and
haemato-immunotoxic7impacts have been reported as the results of TTZ treatment at ADI and below. The main suspect to trigger all these effects is oxidative stress resulting from TTZ
metabolites produced by the gut microbiota8,9. In humans, the most controversial health aspect of TTZ is its association with attention-deficit hyperactivity disorder (ADHD). Dated back to
the 70s10, that allegation once again drew public attention in 2007 thanks to the RCT study of McCann et al.11, listing TTZ among the infamous “Southampton six” – food additives whose
mixture could possibly induce hyperactivity in children. Besides, immune (hypersensitivity) and non-immune (intolerance) reactions to the dye have been long reported, albeit rare3,10,12,13.
One notable TTZ-induced symptom in several cases is the occurrence of purpuric spots (subcutaneous hemorrhage)13,14,15. While most of the studies until now were based on cell culture and
rodent models, zebrafish embryo is increasingly used for toxicological investigations16. Generally considered as a non-regulated embryonic in vivo model (until 120 h old)17, this embryo can
be used in complement to conventional in vitro and animal models: its transparent body allows visualization of all standard vertebrate systems and organs; its rapid development allows
assessment of multiple toxicological targets and processes within a few days; more importantly, research using zebrafish embryos can provide predictive value not only for human health risk
but also for ecotoxicity assessment16,18,19. Here, by combining tests on zebrafish embryos and human cells, we show that TTZ displays a disruptive effect on vascular formation, which may
help shed light on the possible processes involved in some of TTZ’s adverse effects. METHODS MATERIALS The zebrafish wild type (WT) AB strain, and transgenic strains _Tg(fli1:EGFP) (y1Tg)_20
and _Tg(kdrl:mCherry)_(y171Tg)21were maintained within the Zebrafish Facility in GIGA-Research, University of Liège. Fish were reared in a Techniplast (Buguggiate, Italy) recirculating
system under a 14:10-h light/dark photocycle. The isolation and maintenance of human umbilical vein endothelial cells (HUVECs) were previously described22,23. The cells were cultured in EGM2
(Lonza, Verviers, Belgium) supplemented with 5% FCS (Gibco, Thermo Fisher Scientific, Waltham, MA, USA). Tartrazine (#86310, Sigma Aldrich, Hoeilaart, Belgium) 100 g/L stock solution was
prepared and diluted as appropriate in E3 (5 mM NaCl, 0.17 mM KCl, 0.4 mM CaCl2, and 0.16 mM MgSO4) for zebrafish experiments and in PBS for cell experiments. ZEBRAFISH EMBRYONIC ASSAYS
_General toxicity evaluation _The zebrafish embryonic toxicity (ZET) assay was adapted from the OECD Test No.236 (Fish Embryo Toxicity Test)19. After breeding, eggs were collected into E3
medium. At 3–4 h post-fertilization (hpf), fertilized and healthy embryos were selected and distributed into 6-well plates at 25 embryos/well containing 5 mL of E3 medium supplemented or not
with TTZ. Embryos were raised until four days post-fertilization (4 dpf), solutions were renewed and dead embryos were removed daily. Lethality and malformation rates were recorded.
Phenotypes were compared with those described previously by Kimmel et al.24 _Blood vessel assessment_: Transgenic embryos were obtained by crossing heterozygous _Tg(fli1:EGFP)_ or
_Tg(kdrl:mCherry)_ with WT and exposed to 5 and 10 g/L TTZ from 3-4hpf. Visual analysis was performed at 3 dpf using a Leica (Wetzlar, Germany) M165FC fluorescent dissecting microscope. Both
transgenic lines are vasculogenic reporters whose endothelial cells (ECs) are labeled with respectively green or red fluorescence. Despite displaying a brighter fluorescence signal, the
_Tg(fli1:EGFP)_ was used solely for the sub-intestinal vein (SIV) assessment due to the intense fluorescence in the pharyngeal arch cranial neural crest cells. Therefore, the pan-vasculature
observation was performed on the _Tg(kdrl:mCherry)_ line. All experiments were performed in triplicate, _n_ = 50. HUMAN UMBILICAL VEIN ENDOTHELIAL CELL (HUVEC) ASSAYS Human umbilical vein
endothelial cells were obtained from Lonza (Verviers, Belgium) and used between passages 3 and 8. All experiments were performed in triplicate, and each included five wells per TTZ
concentration (including control), each time cell viability was ensured by staining with trypan blue before seeding. _Cell proliferation_ Cells were seeded into a 96-well plate for 48 h at
8000 cells/well. Selected concentrations of TTZ (0–1000 ug/mL) were then added, and proliferation was analyzed after 24 h using the Cell Proliferation ELISA BrdU Colorimetric kit (Roche,
Mannheim, Germany) following the manufacturer’s instructions. Absorbance at 355 nm was measured on a VICTOR X3 Multilabel Reader (PerkinElmer, Waltham, USA). _Tubulogenesis_ HUVECs were
seeded onto a Matrigel-coated 96-well plate at 3000 cells/well into 100 µl of medium supplemented with selected concentrations of TTZ (0–1000 µg/ml), then incubated at 37 °C for 6 h.
Tubulogenic pictures were taken under a CKX41 microscope (Olympus, Tokyo, Japan) and analyzed using ImageJ. _Migration_ HUVECs were plated into 48-well plates and cultured to confluency in
EGM2. A scratch wound was then created in the monolayers using a pipette tip. The scratched wells were washed with PBS and incubated in EBM2 0.5% DBS for 6 h. The wound size (i.e., the
distance between two sides of the wound) was measured at time points 0 h and 6 h with a CKX41 microscope (Olympus, Tokyo, Japan). The difference between wound sizes at 6 h and 0 h (i.e.,
wound closure) was calculated and represented as relative to the respective control per replicate. TARGETNET PREDICTION The online platform TargetNet (http://targetnet.scbdd.com, accessed
January 2022) was used to predict potential protein targets of TTZ using its molecular fingerprint. TTZ’s SMILES string was obtained from PubChem (https://pubchem.ncbi.nlm.nih.gov).
TargetNet parameters were set at default values (AUC ≥ 0.7; Fingerprint type ECFP4). Potential target hits were defined as having a probability score > 0. DATA AND STATISTICAL ANALYSIS
All data analysis and graphs were performed using R 4.3.2. All scripts and raw data are available upon request from the authors. Concerning zebrafish embryo development assessment, toxicity
indices (i.e. LC50 and EC50) of TTZ were estimated by fitting dose response models with two-parameter log-logistic functions using package “drc”25. The teratogenic index (TI) was then
calculated by dividing the LC50 by EC50. Mixed effect models (using package “lme4”26) were fitted to analyze the observed survival and malformation rates at different treatment conditions
due to the multilevel nature of the experiments (including both biological and technical replications). The average survival and malformation rates and their 95% prediction intervals (error
bars) were obtained using the function _predictInterval()_in packages “merTools”27 and then plotted using Prism. A similar approach was also employed for ectopic sprouting data. The same
analytical pipeline and R packages (“drc”, “lme4”, and “merTools”) were used for the results from HUVEC cell experiments. The IC50 of TTZ on HUVEC proliferation was determined using the same
method as the ZET assay. For tube formation data, since both tube length and branching results are bounded by zero and not normally distributed, they were first log-transformed before
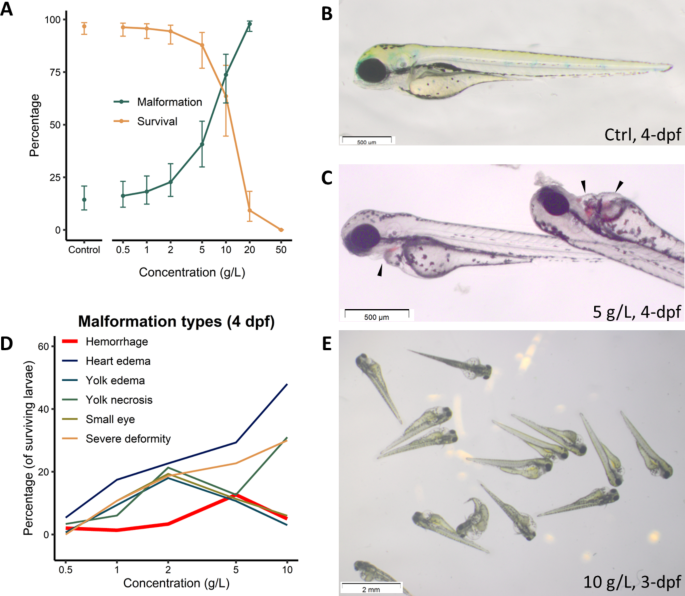
analyzing and model fitting. RESULTS IN VIVO EFFECT OF TTZ ON THE DEVELOPING ZEBRAFISH EMBRYOS HEMORRHAGE AS A REMARKABLE MALFORMATION Following a preliminary range-finding experiment, wild
type zebrafish larvae were exposed to TTZ concentrations ranging from 0 to 50 g/L in order to identify toxicity indices (including LC50, EC50, and TI - teratogenic index) as well as the
types of malformations caused by TTZ exposure. Compared to controls, the test revealed TTZ to be non-aquatoxic with an LC5096hpf of 9.31 g/L and an EC5096hpf of 5.49 g/L (Fig. 1A) (more
description below). Thus, with a TI (teratogenic index, defined as the ratio between LC50 and EC50) of 1.7, the compound can be considered slightly teratogenic for zebrafish embryonic
development (Fig. 1A). Compared to the control (Fig. 1B), one remarkable malformation caused by TTZ exposure was the occurrence of multiple hemorrhagic sites across the embryonic body,
starting at 2 dpf. The severity of hemorrhage was dose-dependent: at lower concentrations, the sites mainly appeared around the duct of Cuvier (i.e., the common cardinal vein) region (Fig.
1C) and then expanded to other body parts at higher concentrations, including brain hemorrhage. Other notable defects included pericardial edema, yolk edema, yolk necrosis, small eye, and
whole-body deformities (Fig. 1D-E). However, the percentage of TTZ-induced bleeding larvae did not follow the typical dose-response relationship (Fig. 1D, thick red line). The decrease in
the percentage of larvae with specific defects (hemorrhage, small eye, yolk edema and necrosis, Fig. 1D) at high concentrations suggests that these treatments lead to multiple, more severe
defects16. Also, the biphasic curves of these specific defects, and the transitions of dominant phenotypes, suggest multiple modes of action involved16. The hemorrhage phenotype, which
reached 14% at 5 g/L then declined at 10 g/L, was genuinely remarkable, thus follow-up investigations were conducted. TTZ TREATMENT OF TRANSGENIC REPORTER FISH LINES REVEALED IMPAIRMENT OF
IN VIVO ANGIOGENESIS Upon discovering the TTZ-induced hemorrhagic phenotype in wild type zebrafish embryos, further experiments were conducted using transgenic reporter zebrafish lines to
verify whether TTZ could affect blood vessel formation. The exposure doses were selected at 5 and 10 g/L as these concentrations produced the most hemorrhagic phenotypes without inducing too
much mortality or other deformities (Fig. 1A, E). Firstly, whole vascular observation was conducted on the _Tg(kdrl:mCherry)_ embryos at 3 dpf. The results revealed that TTZ exposure could
disrupt endothelial cell (EC) organization into the vascular system to various extents in the larvae. The effect was more evident at 10 g/L TTZ, where many larvae had impaired branching and
segmentation in both the cephalic vasculature and the caudal vein plexus (CVP) (Fig. 2A). Notably, the dorsal lateral anastomosis vessels (DLAV) and intersegmental vessels (ISV) are only
marginally affected. Next, to quantify the inhibitory effect of TTZ on the zebrafish blood vessel formation, further experiments were conducted with emphasis on SIV formation18,28 using the
_Tg(fli1:GFP)_. At 3 dpf, a normally developed SIV forms a smooth, basket-shaped structure on each side of the embryonic yolk (Fig. 2B). In our experiments, TTZ exposure at 5 and 10 g/L
respectively induced a smaller basket size and abnormal SIV ectopic sprouting in 50% and 14% of 3-dpf embryos (Fig. 2C) – following the same trend of hemorrhage occurrences at 4-dpf larvae
(Fig. 1E) but at higher percentage. TTZ INHIBITS HUVEC TUBULOGENESIS, MIGRATION, AND PROLIFERATION After confirming the vascular disrupting potential of TTZ on in vivo zebrafish embryos, the
subsequent step is to verify if it was due to the direct effect of the food dye on ECs or possibly another indirect mechanism. Furthermore, to verify whether these effects would be observed
in human cells, several in vitro experiments were carried out using primary Human Umbilical Vascular Endothelial Cells (HUVECs). BrdU incorporation assays revealed that TTZ could inhibit
HUVEC proliferation with an IC50 of 30 mg/L (56 µM) (Fig. 3A). Tubulogenesis assays showed that the compound also inhibited HUVEC tube formation (Fig. 3B-D). Practically, tubes and branches
stopped forming at 267 mg/L (500 µM) (Fig. 3C-D). Additionally, wound healing assays (Fig. 3E-F) revealed that HUVEC cell migration was substantially inhibited by TTZ. All tested effects of
TTZ on HUVEC activities (proliferation, tubulogenesis, and migration) were dose-dependent, while statistical analyses indicated that the observed inhibition was significant even at the
lowest tested dose (10.7 mg/L or 20 µM, _p_ < 0.005). DISCUSSION TTZ AFFECTS BLOOD VESSEL FORMATION, POSSIBLY THROUGH INTERACTION WITH THE BMP SIGNALING PATHWAY Throughout decades of
research on the food dye tartrazine with numerous investigations, this is the first study reporting that TTZ has vascular disrupting properties. We have shown that the dye can impair
angiogenesis by disrupting blood vessel formation in developing zebrafish embryos and by inhibiting human primary ECs from proliferating, migrating, and forming tubular networks using an
array of simple experiments. Combining tests using zebrafish embryos and mammalian ECs has previously been used as a cost-effective tool in studying angiogenic factors29,30,31,32, here it
helped reveal that the vascular disrupting effect of TTZ is universally conserved across multiple species, possibly pan-vertebrate. In zebrafish, the formation of both the DLAV and the ISV
are mainly regulated by the vascular endothelial growth factor (VEGF) signaling pathway (Vegf-A and Vegf-C)33. On the other hand, SIV development is dependent on both the bone morphogenic
protein (BMP) and the VEGF pathway33,34,35,36, while brain vascularization37and EC migration during CVP formation require mainly BMP signaling34,38. Thus, the effects of TTZ observed on
zebrafish embryos predominantly on SIV, CVP, and cranial vessels, while DLAV and ISV are unaffected (Table 1), strongly suggest that this compound may affect vascularization by interacting
with the BMP pathway, rather than the more popular VEGF-A signaling. RHO GTPASES AS THE MAIN SUSPECT The action of BMP signaling in zebrafish blood vessel formation is closely linked with
activities of the Rho GTPase family, which regulates cell migration by controlling the formation of cytoskeleton processes32,39,40,41. Wakayama et al. (2015) reported BMP action mediated by
Cdc42, a Rho GTPase family member whose inhibition leads to a fused CVP network without clear spaces between capillaries34. Cdc42 has been shown to control EC directional sensing through
projection of filopodia39, whose disruption could lead to inhibition of the wound healing process31. It also regulates microtubule alignment and EC elongation29,42; hence its
loss-of-function would facilitate EC lumen collapse, which, together with the impaired migration, could further contribute to tubulogenic failure. Another family member is RAC1, which plays
multiple roles in endothelial cell activities and vascular development43, contributing to EC survival41, migration39, and zebrafish brain vascular pruning44. Interestingly, human RAC1
protein was predicted to interact with TTZ by the TargetNet platform (Supplementary Table S1). The third family member RHOA also controls EC directional migration45,46and tube collapse47. A
recent paper also suggested key roles of Rac1 and RhoA in the zebrafish BMP angiogenic pathways32. While no direct relationship between Rho GTPases and zebrafish SIV ectopic formation was
found in the literature, indirect evidence exists, such as via elements of the lipid metabolism pathways48– for example, treatment with the cholesterol-reducing drug atorvastatin led to both
SIV ectopic sprouting in zebrafish49and an increase of GTP loading of all Rho GTPases in HUVECs50. Another indirect evidence is the family of cerebral cavernous malformations genes (CCMs),
whose pathways regulate Rho GTPases in ECs51,52. Loss-of-function of CCM orthologs in zebrafish causes ectopic SIV sprouting and fused CVP53,54, and induce diverse effects on human EC
proliferation, migration, and tube formation depending on both EC type (HUVEC or brain EC)55and the CCM gene involved (CCM1, 2, or 3)52,55,56. Noteworthily, variants of the CCM genes are
also responsible for many cases of brain hemorrhage51,52,54,55. On the other hand, TTZ treatment in vitro (in human hepatoma cells)57and in vivo (in CD1 mice)58led to changes in expression
of several genes participating in RhoA GTPase signaling pathways: JUN, MAPK8 (JNK1)59,60, MAP2K6 (MKK6)60,61, and PIK3C2A62,63. Re-analyzing the transcriptomic changes following TTZ
treatment in human hepatoma cells57 further revealed regulation of genes related to Rho GTPase-activated formins, Rho GTPase effectors, and signaling by Rho GTPases. In addition to RAC1,
TargetNet also suggests other Rho GTPase signaling-related proteins capable of interacting with TTZ (Supplementary Table S1). They include HDAC4, which could antagonize KLF2-induced RHOA
activation and promote random cell motility64, or PTGS2, which is downstream of RHOA/RAC1/CDC42 in endothelia65and fibroblasts66. These observations further support the possible interactions
between TTZ and Rho family GTPase activities. OTHER POSSIBLE INTERACTIONS Aside from the possibility of interacting with Rho GTPases, TTZ may act via multiple pathways, as the compound was
reported to have both estrogenic67and oxidative68potentials – both of which are related to the angiogenesis pathway69,70and GTPase activities69,71. Indeed, our ZET results corroborate the
involvement of multiple toxicological mechanisms beside vascularization (Fig. 1E). Still, further investigations are required to comprehensively understand TTZ’s vascular disruptive effect
and other reported bioactivities. In this study, one striking observation is the difference in effective concentrations of TTZ in the two systems: in zebrafish larvae, substantial effects on
vascular development are seen at 5–10 g/L, while HUVECs display decreased activities at 10.7 mg/L (20 µM) and completely malfunction at 267 mg/L (0.5 mM). It is important to note here that
TTZ is a hydrophilic compound, dissolved in water, thus the chorionic or other cell membranes may protect the larvae from its influence, while HUVECs are directly exposed to it. In addition,
the detoxifying organs (i.e., liver and kidney) may be involved, as well as the affinity of serum albumin to TTZ1,3. While the exact mechanism of TTZ’s vascular disrupting feature is yet to
be determined, our discovery may already offer several applied implications. Tartrazine is a common, yet controversial food additive that is alleged to cause multiple adverse effects,
including genotoxicity, oxidative stress, and behavioral alteration1,2,4,6,7,8,9,10,12,13,15,67,68,72. However, no report has mentioned its effect on the vascular system or endothelium, even
though hemorrhagic sites can be observed in previous studies73,74. This newly found anti-angiogenic feature may shed light on the presence of purpura experienced by some individuals14,15,
which has long been attributed to an immune response12,14,15. Our discovery may offer a straightforward explanation for these cases: ECs exposed to TTZ will malfunction, leading to vascular
defect and internal hemorrhage. The reason why TTZ-induced purpura rarely happens could be explained by either a high individual sensitivity to its effects, or the fact that more often TTZ
does not reach an effective concentration within the blood vessels: less than 5% of TTZ is absorbed after oral uptake, then predominantly excreted via urine1, whereas a large proportion of
the chemical may have been metabolized by the gut microflora2. Regarding the emerging recognition of gut microbiota in food allergy cases75, it is reasonable to hypothesize a link between
the microbiota and TTZ hypersensitivity. Another health aspect of TTZ, perhaps the most controversial, is its alleged link with ADHD in children3,11. If the link is valid, one possible
explanation is the indirect impact of TTZ consumption on brain function via blood vessel function76. The discovered vascular effect of TTZ may also contribute to the safety regulation of the
colorant, especially in food and medical applications. Methods have been published using TTZ as a photoabsorber for light-based hydrogel bioprinting to be used as scaffolds for endothelial
cell seeding and tissue engineering77,78,79,80. Even though there is a washing step between scaffold printing and cell seeding, the risk of TTZ residues affecting HUVECs behaviors should not
be neglected. More importantly, TTZ as a common drug excipient and food additive may interfere with proangiogenic therapy such as in coronary and peripheral artery disease or ischemic
stroke81. On the other hand, this TTZ’s function could lead to potential applications in treating conditions such as cancer or macular degeneration28,81,82. Given the extensive information
on TTZ toxicity and safety, its status as a food additive, and its abundant and cheap production, the chemical may serve as an affordable alternative for existing anti-angiogenic drugs. DATA
AVAILABILITY All scripts and raw data are available upon request from the authors - namely M.M., D.D.T., and N.B.N. REFERENCES * EFSA. Scientific opinion on the re-evaluation tartrazine (E
102). _EFSA J._ 7, 1331. https://doi.org/10.2903/j.efsa.2009.1331 (2009). Article Google Scholar * EFSA. Statement on Allura Red AC and other sulphonated mono azo dyes authorised as food
and feed additives. _EFSA J._ 11, 3234. https://doi.org/10.2903/j.efsa.2013.3234 (2013). Article Google Scholar * JECFA. _Safety Evaluation of Certain food Additives: Prepared by the
Eighty-second Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA)_v, 493 (p. (World Health Organization, 2017). Google Scholar * Khayyat, L., Essawy, A., Sorour, J.
& Soffar, A. Tartrazine induces structural and functional aberrations and genotoxic effects in vivo. _PeerJ_ 5, e3041. https://doi.org/10.7717/peerj.3041 (2017). Article CAS PubMed
PubMed Central Google Scholar * Hashem, M. M., Abd-Elhakim, Y. M., Abo-El-Sooud, K. & Eleiwa, M. M. E. Embryotoxic and Teratogenic effects of Tartrazine in rats. _Toxicol. Res._ 35,
75–81. https://doi.org/10.5487/tr.2019.35.1.075 (2019). Article CAS PubMed Google Scholar * Bhatt, D., Vyas, K., Singh, S., John, P. J. & Soni, I. Tartrazine induced neurobiochemical
alterations in rat brain sub-regions. _Food Chem. Toxicol._ 113, 322–327. https://doi.org/10.1016/j.fct.2018.02.011 (2018). Article CAS PubMed Google Scholar * Abd-Elhakim, Y. M. et al.
Comparative haemato-immunotoxic impacts of long-term exposure to tartrazine and chlorophyll in rats. _Int. Immunopharmacol._ 63, 145–154. https://doi.org/10.1016/j.intimp.2018.08.002
(2018). Article CAS PubMed Google Scholar * Gao, Y. et al. Effect of food azo dye tartrazine on learning and memory functions in mice and rats, and the possible mechanisms involved. _J.
Food Sci._ 76, T125–129. https://doi.org/10.1111/j.1750-3841.2011.02267.x (2011). Article CAS PubMed Google Scholar * Himri, I. et al. A 90-day oral toxicity study of Tartrazine, a
synthetic food dye, in Wistar rats. _Int. J. Pharm. Pharm. Sci._ 3, 159–169 (2011). CAS Google Scholar * Simon, R. A. Adverse reactions to food additives. _Curr. Allergy Asthma Rep._ 3,
62–66. https://doi.org/10.1007/s11882-003-0014-9 (2003). Article PubMed Google Scholar * McCann, D. et al. Food additives and hyperactive behaviour in 3-year-old and 8/9-year-old children
in the community: a randomised, double-blinded, placebo-controlled trial. _Lancet_ 370, 1560–1567. https://doi.org/10.1016/S0140-6736(07)61306-3 (2007). https://doi.org/http://dx. Article
CAS PubMed Google Scholar * Miller, K. Sensitivity to tartrazine. _Br. Med. J. (Clin Res. Ed)_. 285, 1597–1598 (1982). Article CAS PubMed Google Scholar * Gultekin, F. & Doguc, D.
K. Allergic and immunologic reactions to food additives. _Clin. Rev. Allergy Immunol._ 45, 6–29. https://doi.org/10.1007/s12016-012-8300-8 (2013). Article CAS PubMed Google Scholar *
Criep, L. H. Allergic vascular purpura. _J. Allergy Clin. Immunol._ 48, 7–12. https://doi.org/10.1016/0091-6749(71)90049-2 (1971). https://doi.org/http://dx. Article CAS PubMed Google
Scholar * Parodi, G., Parodi, A. & Rebora, A. Purpuric Vasculitis due to Tartrazine. _Dermatology_ 171, 62–63 (1985). Article CAS Google Scholar * Babayigit, A. et al. Assessing the
toxicity of Pb- and Sn-based perovskite solar cells in model organism Danio rerio. _Sci. Rep._ 6, 18721. https://doi.org/10.1038/srep18721 (2016). Article ADS CAS PubMed PubMed Central
Google Scholar * European Council. (2010). * Serbedzija, G. N., Flynn, E. & Willett, C. E. Zebrafish angiogenesis: a new model for drug screening. _Angiogenesis_ 3, 353–359.
https://doi.org/10.1023/a:1026598300052 (1999). Article CAS PubMed Google Scholar * OECD. _Test No. 236: Fish Embryo Acute Toxicity (FET) Test_ (OECD Publishing, 2013). Book Google
Scholar * Lawson, N. D. & Weinstein, B. M. In vivo imaging of embryonic vascular development using transgenic zebrafish. _Dev. Biol._ 248, 307–318.
https://doi.org/10.1006/dbio.2002.0711 (2002). Article CAS PubMed Google Scholar * Fujita, M. et al. Assembly and patterning of the vascular network of the vertebrate hindbrain.
_Development_ 138, 1705–1715. https://doi.org/10.1242/dev.058776 (2011). Article CAS PubMed PubMed Central Google Scholar * Jaffe, E. A., Nachman, R. L., Becker, C. G. & Minick, C.
R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. _J. Clin. Invest._ 52, 2745–2756.
https://doi.org/10.1172/JCI107470 (1973). Article CAS PubMed PubMed Central Google Scholar * Fontaine, M. et al. Extracellular vesicles mediate communication between endothelial and
vascular smooth muscle cells. _Int. J. Mol. Sci._ 23 https://doi.org/10.3390/ijms23010331 (2021). * Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B. & Schilling, T. F. Stages of
embryonic development of the zebrafish. _Dev. Dyn._ 203, 253–310. https://doi.org/10.1002/aja.1002030302 (1995). Article CAS PubMed Google Scholar * Ritz, C., Baty, F., Streibig, J. C.
& Gerhard, D. Dose-response analysis using R. _PLoS One_. 10, e0146021. https://doi.org/10.1371/journal.pone.0146021 (2015). Article CAS PubMed PubMed Central Google Scholar *
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting Linear mixed-effects models Usinglme4. _J. Stat. Softw._ 67 https://doi.org/10.18637/jss.v067.i01 (2015). * Knowles, J. &
Frederick, C. _merTools: Tools for Analyzing Mixed Effect Regression Models_, < (2020). https://CRAN.R-project.org/package=merTools * Chimote, G. et al. Comparison of effects of
anti-angiogenic agents in the zebrafish efficacy-toxicity model for translational anti-angiogenic drug discovery. _Drug Des. Devel Ther._ 8, 1107–1123. https://doi.org/10.2147/DDDT.S55621
(2014). Article CAS PubMed PubMed Central Google Scholar * Hetheridge, C. et al. The formin FMNL3 is a cytoskeletal regulator of angiogenesis. _J. Cell. Sci._ 125, 1420–1428.
https://doi.org/10.1242/jcs.091066 (2012). Article CAS PubMed PubMed Central Google Scholar * McCollum, C. W. et al. Identification of vascular disruptor compounds by analysis in
zebrafish embryos and mouse embryonic endothelial cells. _Reprod. Toxicol._ 70, 60–69. https://doi.org/10.1016/j.reprotox.2016.11.005 (2017). Article CAS PubMed Google Scholar * Lavina,
B. et al. Defective endothelial cell migration in the absence of Cdc42 leads to capillary-venous malformations. _Development_ 145 https://doi.org/10.1242/dev.161182 (2018). * Veloso, A. et
al. The cytoskeleton adaptor protein Sorbs1 controls the development of lymphatic and venous vessels in zebrafish. _BMC Biol._ 22, 51. https://doi.org/10.1186/s12915-024-01850-z (2024).
Article CAS PubMed PubMed Central Google Scholar * Nakajima, H., Chiba, A., Fukumoto, M., Morooka, N. & Mochizuki, N. Zebrafish Vascular Development: General and tissue-specific
regulation. _J. Lipid Atheroscler_. 10, 145–159. https://doi.org/10.12997/jla.2021.10.2.145 (2021). Article CAS PubMed PubMed Central Google Scholar * Wakayama, Y., Fukuhara, S., Ando,
K., Matsuda, M. & Mochizuki, N. Cdc42 mediates Bmp-induced sprouting angiogenesis through Fmnl3-driven assembly of endothelial filopodia in zebrafish. _Dev. Cell._ 32, 109–122.
https://doi.org/10.1016/j.devcel.2014.11.024 (2015). Article CAS PubMed Google Scholar * Goi, M. & Childs, S. J. Patterning mechanisms of the sub-intestinal venous plexus in
zebrafish. _Dev. Biol._ 409, 114–128. https://doi.org/10.1016/j.ydbio.2015.10.017 (2016). Article CAS PubMed Google Scholar * Koenig, A. L. et al. Vegfa signaling promotes zebrafish
intestinal vasculature development through endothelial cell migration from the posterior cardinal vein. _Dev. Biol._ 411, 115–127. https://doi.org/10.1016/j.ydbio.2016.01.002 (2016). Article
CAS PubMed PubMed Central Google Scholar * Rochon, E. R., Menon, P. G. & Roman, B. L. Alk1 controls arterial endothelial cell migration in lumenized vessels. _Development_ 143,
2593–2602. https://doi.org/10.1242/dev.135392 (2016). Article CAS PubMed PubMed Central Google Scholar * Phng, L. K., Stanchi, F. & Gerhardt, H. Filopodia are dispensable for
endothelial tip cell guidance. _Development_ 140, 4031–4040. https://doi.org/10.1242/dev.097352 (2013). Article CAS PubMed Google Scholar * Lamalice, L., Le Boeuf, F. & Huot, J.
Endothelial cell migration during angiogenesis. _Circ. Res._ 100, 782–794. https://doi.org/10.1161/01.RES.0000259593.07661.1e (2007). Article CAS PubMed Google Scholar * van der Meel, R.
et al. The VEGF/Rho GTPase signalling pathway: a promising target for anti-angiogenic/anti-invasion therapy. _Drug Discov Today_. 16, 219–228. https://doi.org/10.1016/j.drudis.2011.01.005
(2011). Article CAS PubMed Google Scholar * Boueid, M. J., Mikdache, A., Lesport, E., Degerny, C. & Tawk, M. Rho GTPases signaling in zebrafish development and disease. _Cells_ 9
https://doi.org/10.3390/cells9122634 (2020). * Phng, L. K. et al. Formin-mediated actin polymerization at endothelial junctions is required for vessel lumen formation and stabilization.
_Dev. Cell._ 32, 123–132. https://doi.org/10.1016/j.devcel.2014.11.017 (2015). Article CAS PubMed Google Scholar * Tan, W. et al. An essential role for Rac1 in endothelial cell function
and vascular development. _FASEB J._ 22, 1829–1838. https://doi.org/10.1096/fj.07-096438 (2008). Article CAS PubMed Google Scholar * Chen, Q. et al. Haemodynamics-driven developmental
pruning of brain vasculature in zebrafish. _PLoS Biol._ 10, e1001374. https://doi.org/10.1371/journal.pbio.1001374 (2012). Article CAS PubMed PubMed Central Google Scholar * Vega, F.
M., Fruhwirth, G., Ng, T. & Ridley, A. J. RhoA and RhoC have distinct roles in migration and invasion by acting through different targets. _J. Cell. Biol._ 193, 655–665.
https://doi.org/10.1083/jcb.201011038 (2011). Article CAS PubMed PubMed Central Google Scholar * Wu, C. et al. Rab13-dependent trafficking of RhoA is required for directional migration
and angiogenesis. _J. Biol. Chem._ 286, 23511–23520. https://doi.org/10.1074/jbc.M111.245209 (2011). Article CAS PubMed PubMed Central Google Scholar * Bayless, K. J. & Davis, G. E.
Microtubule depolymerization rapidly collapses capillary tube networks in vitro and angiogenic vessels in vivo through the small GTPase rho. _J. Biol. Chem._ 279, 11686–11695.
https://doi.org/10.1074/jbc.M308373200 (2004). Article CAS PubMed Google Scholar * Kalaivani, V. & Jaleel, A. Apolipoprotein(a), an enigmatic anti-angiogenic glycoprotein in human
plasma: a curse or cure? _Pharmacol. Res._ 158, 104858. https://doi.org/10.1016/j.phrs.2020.104858 (2020). Article CAS PubMed Google Scholar * Avraham-Davidi, I. et al. ApoB-containing
lipoproteins regulate angiogenesis by modulating expression of VEGF receptor 1. _Nat. Med._ 18, 967–973. https://doi.org/10.1038/nm.2759 (2012). Article CAS PubMed PubMed Central Google
Scholar * Xiao, H., Qin, X., Ping, D. & Zuo, K. Inhibition of rho and Rac geranylgeranylation by atorvastatin is critical for preservation of endothelial junction integrity. _PLoS One_.
8, e59233. https://doi.org/10.1371/journal.pone.0059233 (2013). Article ADS CAS PubMed PubMed Central Google Scholar * Whitehead, K. J. et al. The cerebral cavernous malformation
signaling pathway promotes vascular integrity via rho GTPases. _Nat. Med._ 15, 177–184. https://doi.org/10.1038/nm.1911 (2009). Article CAS PubMed PubMed Central Google Scholar *
Richardson, B. T., Dibble, C. F., Borikova, A. L. & Johnson, G. L. Cerebral cavernous malformation is a vascular disease associated with activated RhoA signaling. _Biol. Chem._ 394,
35–42. https://doi.org/10.1515/hsz-2012-0243 (2013). Article CAS PubMed PubMed Central Google Scholar * Hogan, B. M., Bussmann, J., Wolburg, H. & Schulte-Merker, S. ccm1 cell
autonomously regulates endothelial cellular morphogenesis and vascular tubulogenesis in zebrafish. _Hum. Mol. Genet._ 17, 2424–2432. https://doi.org/10.1093/hmg/ddn142 (2008). Article CAS
PubMed Google Scholar * Voss, K. et al. Functional analyses of human and zebrafish 18-amino acid in-frame deletion pave the way for domain mapping of the cerebral cavernous malformation 3
protein. _Hum. Mutat._ 30, 1003–1011. https://doi.org/10.1002/humu.20996 (2009). Article CAS PubMed Google Scholar * Zhu, Y. et al. Differential angiogenesis function of CCM2 and CCM3 in
cerebral cavernous malformations. _Neurosurg. Focus_. 29, E1. https://doi.org/10.3171/2010.5.FOCUS1090 (2010). Article PubMed Google Scholar * Zhu, Y. et al. In vitro characterization of
the angiogenic phenotype and genotype of the endothelia derived from sporadic cerebral cavernous malformations. _Neurosurgery_ 69, 722–731. https://doi.org/10.1227/NEU.0b013e318219569f
(2011). discussion 731 – 722. Article PubMed Google Scholar * Gijbels, E., Devisscher, L. & Vinken, M. Testing in vitro tools for the prediction of cholestatic liver injury induced by
non-pharmaceutical chemicals. _Food Chem. Toxicol._ 152, 112165. https://doi.org/10.1016/j.fct.2021.112165 (2021). Article CAS PubMed Google Scholar * Raposa, B. et al. Food additives:
Sodium benzoate, potassium sorbate, azorubine, and tartrazine modify the expression of NFkappaB, GADD45alpha, and MAPK8 genes. _Physiol. Int._ 103, 334–343.
https://doi.org/10.1556/2060.103.2016.3.6 (2016). Article CAS PubMed Google Scholar * Teramoto, H. et al. Signaling from the small GTP-binding proteins Rac1 and Cdc42 to the c-Jun
N-terminal kinase/stress-activated protein kinase pathway. A role for mixed lineage kinase 3/protein-tyrosine kinase 1, a novel member of the mixed lineage kinase family. _J. Biol. Chem._
271, 27225–27228. https://doi.org/10.1074/jbc.271.44.27225 (1996). Article CAS PubMed Google Scholar * Canovas, B. & Nebreda, A. R. Diversity and versatility of p38 kinase signalling
in health and disease. _Nat. Rev. Mol. Cell. Biol._ 22, 346–366. https://doi.org/10.1038/s41580-020-00322-w (2021). Article CAS PubMed PubMed Central Google Scholar * Anton, R., Bauer,
S. M., Keck, P. R., Laufer, S. & Rothbauer, U. A p38 substrate-specific MK2-EGFP translocation assay for identification and validation of new p38 inhibitors in living cells: a
comprising alternative for acquisition of cellular p38 inhibition data. _PLoS One_. 9, e95641. https://doi.org/10.1371/journal.pone.0095641 (2014). Article ADS CAS PubMed PubMed Central
Google Scholar * Yoshioka, K. et al. Endothelial PI3K-C2alpha, a class II PI3K, has an essential role in angiogenesis and vascular barrier function. _Nat. Med._ 18, 1560–1569.
https://doi.org/10.1038/nm.2928 (2012). Article CAS PubMed Google Scholar * Sunitha, P. et al. Temporal VEGFA responsive genes in HUVECs: gene signatures and potential ligands/receptors
fine-tuning angiogenesis. _J. Cell. Commun. Signal._ 13, 561–571. https://doi.org/10.1007/s12079-019-00541-7 (2019). Article CAS PubMed PubMed Central Google Scholar * Cernotta, N.,
Clocchiatti, A., Florean, C. & Brancolini, C. Ubiquitin-dependent degradation of HDAC4, a new regulator of random cell motility. _Mol. Biol. Cell._ 22, 278–289.
https://doi.org/10.1091/mbc.E10-07-0616 (2011). Article CAS PubMed PubMed Central Google Scholar * Schmeck, B. et al. Rho protein inhibition blocks cyclooxygenase-2 expression by
proinflammatory mediators in endothelial cells. _Inflammation_ 27, 89–95. https://doi.org/10.1023/a:1023278600596 (2003). Article CAS PubMed Google Scholar * Chang, Y. W. et al.
Differential regulation of cyclooxygenase 2 expression by small GTPases ras, Rac1, and RhoA. _J. Cell. Biochem._ 96, 314–329. https://doi.org/10.1002/jcb.20568 (2005). Article CAS PubMed
Google Scholar * Axon, A. et al. Tartrazine and sunset yellow are xenoestrogens in a new screening assay to identify modulators of human oestrogen receptor transcriptional activity.
_Toxicology_ 298, 40–51. https://doi.org/10.1016/j.tox.2012.04.014 (2012). Article CAS PubMed Google Scholar * Albasher, G. et al. Perinatal exposure to Tartrazine Triggers Oxidative
Stress and neurobehavioral alterations in mice offspring. _Antioxid. (Basel)_. 9. https://doi.org/10.3390/antiox9010053 (2020). * Moldovan, L., Mythreye, K., Goldschmidt-Clermont, P. J.
& Satterwhite, L. L. Reactive oxygen species in vascular endothelial cell motility. Roles of NAD(P)H oxidase and Rac1. _Cardiovasc. Res._ 71, 236–246.
https://doi.org/10.1016/j.cardiores.2006.05.003 (2006). Article CAS PubMed Google Scholar * Fatima, L. A. et al. Estrogen receptor 1 (ESR1) regulates VEGFA in adipose tissue. _Sci. Rep._
7, 16716. https://doi.org/10.1038/s41598-017-16686-7 (2017). Article ADS CAS PubMed PubMed Central Google Scholar * Briz, V. & Baudry, M. Estrogen Regulates Protein Synthesis and
actin polymerization in hippocampal neurons through different molecular mechanisms. _Front. Endocrinol. (Lausanne)_. 5, 22. https://doi.org/10.3389/fendo.2014.00022 (2014). * Amin, K.,
Hameid, I. I., Abd Elsttar, A. & H. A. & Effect of food azo dyes tartrazine and carmoisine on biochemical parameters related to renal, hepatic function and oxidative stress
biomarkers in young male rats. _Food Chem. Toxicol._ 48, 2994–2999 (2010). Article CAS PubMed Google Scholar * Jiang, L. L. et al. Toxicity Assessment of 4 Azo Dyes in zebrafish embryos.
_Int. J. Toxicol._ 39, 115–123. https://doi.org/10.1177/1091581819898396 (2020). Article CAS PubMed Google Scholar * Gupta, R. et al. Toxic effects of Food colorants Erythrosine and
Tartrazine on zebrafish embryo development. _Curr. Res. Nutr. Food Sci. J._ 7, 876–885. https://doi.org/10.12944/crnfsj.7.3.26 (2019). Article Google Scholar * Shu, S. A. et al. Microbiota
and Food Allergy. _Clin. Rev. Allergy Immunol._ 57, 83–97. https://doi.org/10.1007/s12016-018-8723-y (2019). Article CAS PubMed Google Scholar * Cannon Homaei, S. et al. ADHD symptoms
in neurometabolic diseases: underlying mechanisms and clinical implications. _Neurosci. Biobehav Rev._ 132, 838–856. https://doi.org/10.1016/j.neubiorev.2021.11.012 (2022). Article CAS
PubMed Google Scholar * Benjamin, A. D. et al. Light-based 3D printing of hydrogels with high-resolution channels. _Biomedical Phys. Eng. Express_. 5
https://doi.org/10.1088/2057-1976/aad667 (2019). * Grigoryan, B. et al. Multivascular networks and functional intravascular topologies within biocompatible hydrogels. _Science_ 364, 458–464.
https://doi.org/10.1126/science.aav9750 (2019). Article ADS CAS PubMed PubMed Central Google Scholar * Kinstlinger, I. S. et al. Perfusion and endothelialization of engineered tissues
with patterned vascular networks. _Nat. Protoc._ 16, 3089–3113. https://doi.org/10.1038/s41596-021-00533-1 (2021). Article CAS PubMed Google Scholar * Yu, K. et al. Printability during
projection-based 3D bioprinting. _Bioact Mater._ 11, 254–267. https://doi.org/10.1016/j.bioactmat.2021.09.021 (2022). Article CAS PubMed Google Scholar * Rust, R., Gantner, C. &
Schwab, M. E. Pro- and antiangiogenic therapies: current status and clinical implications. _FASEB J._ 33, 34–48. https://doi.org/10.1096/fj.201800640RR (2019). Article CAS PubMed Google
Scholar * Fallah, A. et al. Therapeutic targeting of angiogenesis molecular pathways in angiogenesis-dependent diseases. _Biomed. Pharmacother_. 110, 775–785.
https://doi.org/10.1016/j.biopha.2018.12.022 (2019). Article CAS PubMed Google Scholar Download references ACKNOWLEDGEMENTS The authors would like to thank the GIGA zebrafish facility
for providing zebrafish adults for spawning and the GIGA imaging platform for their help and support with microscopy. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Lab. for Organogenesis and
Regeneration, GIGA-Institute, Université de Liège, Liège, 4000, Belgium Dinh Duy Thanh & Marc Muller * Department of Cell Biology, Faculty of Biology, VNU University of Science, Hanoi,
100000, Vietnam Dinh Duy Thanh * VNU School of Interdisciplinary Sciences and Arts, Vietnam National University, Hanoi, 100000, Vietnam Nguyen Bich-Ngoc * Lab. of Molecular Angiogenesis,
GIGA-Institute, Université de Liège, Liège, 4000, Belgium Cécile Paques, Aurélie Christian, Stéphanie Herkenne & Ingrid Struman Authors * Dinh Duy Thanh View author publications You can
also search for this author inPubMed Google Scholar * Nguyen Bich-Ngoc View author publications You can also search for this author inPubMed Google Scholar * Cécile Paques View author
publications You can also search for this author inPubMed Google Scholar * Aurélie Christian View author publications You can also search for this author inPubMed Google Scholar * Stéphanie
Herkenne View author publications You can also search for this author inPubMed Google Scholar * Ingrid Struman View author publications You can also search for this author inPubMed Google
Scholar * Marc Muller View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS D.D.T. and M.M. conceived the study; D.D.T. carried out the zebrafish
experiments; N.B.N. performed data analysis and visualization; C.P., A.C., S.H., and I.S. carried out the HUVEC experiments; D.D.T., M.M., N.B.N., S.H., and I.S. contributed to writing the
paper. CORRESPONDING AUTHOR Correspondence to Marc Muller. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL Below is the link to the electronic
supplementary material. SUPPLEMENTARY MATERIAL 1 RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0
International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s)
and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material
derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a
credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted
use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/. Reprints and permissions
ABOUT THIS ARTICLE CITE THIS ARTICLE Thanh, D.D., Bich-Ngoc, N., Paques, C. _et al._ The food dye Tartrazine disrupts vascular formation both in zebrafish larvae and in human primary
endothelial cells. _Sci Rep_ 14, 30367 (2024). https://doi.org/10.1038/s41598-024-82076-5 Download citation * Received: 21 July 2024 * Accepted: 02 December 2024 * Published: 05 December
2024 * DOI: https://doi.org/10.1038/s41598-024-82076-5 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable
link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * Tartrazine * Blood vessels, Zebrafish *
HUVEC * BMP pathways * Rho GTPases