Play all audios:
ABSTRACT Exposure to excess glucocorticoid (GC) during early development is implicated in adult dysfunctions. Reduced adult hippocampal neurogenesis is a well-known consequence of exposure
to early life stress or elevated GC, however the effects on neurogenesis during development and effects on other brain regions are not well understood. Using an optogenetic zebrafish model,
here we analyse the effects of GC exposure on neurogenesis during development in the whole brain. We identify that the hypothalamus is a highly GC-sensitive region where elevated GC causes
precocious development. This is followed by failed maturation and early decline accompanied by impaired feeding, growth, and survival. In GC-exposed animals, the developmental trajectory of
hypothalamic progenitor cells is strikingly altered, potentially mediated by direct regulation of transcription factors such as _rx3_ by GC. Our data provide cellular and molecular level
insight into GC-induced alteration of the hypothalamic developmental trajectory, a process crucial for health across the life-course. SIMILAR CONTENT BEING VIEWED BY OTHERS LONG LASTING
ANXIETY FOLLOWING EARLY LIFE STRESS IS DEPENDENT ON GLUCOCORTICOID SIGNALING IN ZEBRAFISH Article Open access 27 July 2022 GESTATIONAL BISPHENOL A EXPOSURE ALTERS ENERGY HOMEOSTASIS AND
ADULT HYPOTHALAMIC NEUROGENESIS IN FEMALE MICE Article Open access 12 July 2024 PRENATAL EXPOSURE TO THE MINERALOCORTICOID RECEPTOR ANTAGONIST SPIRONOLACTONE DISRUPTS HIPPOCAMPAL AREA CA2
CONNECTIVITY AND ALTERS BEHAVIOR IN MICE Article Open access 05 September 2024 INTRODUCTION Glucocorticoids (GC) are the key effectors of the stress response and have pleiotropic effects on
the body, acting to restore homoeostasis and thus allow an animal to respond adaptively to threat1. Since the developing brain is plastic, GC exposure during early life has the potential to
alter the developmental trajectory of the brain. Indeed, alteration of the developmental trajectory may be an adaptive mechanism employed by animals exposed to stress during early life. For
example, antenatal GC treatment has well documented effects on lung maturation, as well as other organs, in preterm babies2. However, such treatment has also been associated with negative
outcomes in later life, such as increased incidence of mental health issues during childhood3, impacted development of fronto-parietal brain functions during adolescence4, and cortical
thinning in children with associated affective disorders5. Early life GC exposure leading to adverse consequences in later life is an example of the concept known as early life programming
of adult disease6,7,8,9. One of the well-documented effects of GC exposure during early life or early life stress (ELS) is reduced adult neurogenesis in the hippocampus, including reduced
cell proliferation10,11,12,13,14. Indeed, adrenalectomy is sufficient to prevent reduced proliferation in chronically stressed mice, suggesting that GC is the main driving force behind this
phenotype15. In contrast, only a few studies have looked at the effects of ELS or GC exposure on neurogenesis during development, but there are some indications that its effects might be
developmentally dynamic. Specifically, it is possible that GC-induced reduction of cell proliferation is a delayed consequence of a GC-induced change to the developmental trajectory that
began in early life. Indeed, rats exposed to ELS exhibited enhanced hippocampal cell proliferation and improved stress-associated behavioural performance as young adults, whilst these
effects were reversed by middle age10. A potential mechanistic explanation for the ELS-induced reduction in adult hippocampal neurogenesis is that enhanced cell proliferation at an earlier
time-point depletes the stem cell pool over time16. Despite the well-documented effects of GC exposure on hippocampal neurogenesis, the effects of GC exposure on other brain regions are
unclear. A recent study revealed the effects of ELS on hypothalamic neurogenesis showing that cell proliferation and numbers of hypothalamic stem cells known as tanycytes were reduced in the
adult mouse17. Tanycytes are unique radial glial-like cells that line the walls of the 3rd ventricle, have NSPC (neural stem/progenitor cell) properties that persist into adulthood, send
projections into neighbouring hypothalamic nuclei and are able to sense glucose levels in the CSF18. Whilst most previous studies have focused on effects of stress and GC exposure on
neurogenesis in the hippocampus, the aforementioned study supports that the hippocampus is not uniquely affected by stress/GC. However, whether and how GC affects hypothalamic neurogenesis
during development is largely unknown. Here we used an optogenetic zebrafish model in which the endogenous cortisol level is elevated, to test the hypothesis that GC exposure alters the
developmental trajectory of the brain. The model utilises optogenetic manipulation of steroidogenic interrenal cells to increase the endogenous GC level during development19,20,21. Whilst
the mammalian adrenal gland, counterpart of the fish interrenal, is known to produce GCs, mineralocorticoids (MCs), and androgens22; MCs and androgens are not detectable in the larval
zebrafish interrenal gland23,24,25,26. Therefore, the phenotypes observed in our model are primarily caused by GC and its downstream effectors, rather than MC or androgen over-exposure. We
found that GC exposure affected cell proliferation in a region-specific manner and that these effects were dynamic across the life-course. We found that the hypothalamus is a highly
GC-sensitive brain region. In early life, we observed precocious development of the hypothalamus and hypothalamus-associated feeding behaviour in GC-exposed animals. This was followed by a
rapid decline, indicated by failed hypothalamic maturation, including reduced cell proliferation and altered numbers of hypothalamic neuronal subtypes, and suppressed feeding and growth. Our
data uncover a tissue-specific time-dependent plasticity of the hypothalamus in response to GC and provide cellular and molecular insights for GC-induced alteration of the hypothalamic
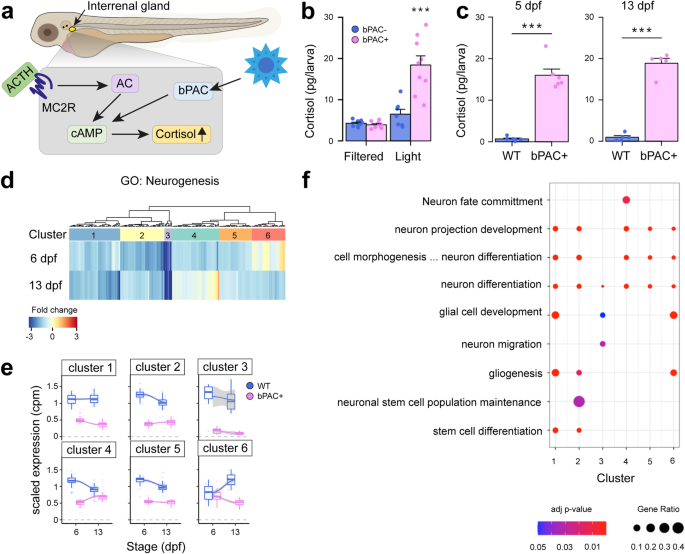
developmental trajectory. RESULTS NEUROGENESIS IS ALTERED IN AN OPTOGENETIC ZEBRAFISH MODEL OF ELEVATED GC We previously developed an optogenetic zebrafish line to induce elevated endogenous
cortisol levels through _beggiatoa_ Photoactivated Adenylate Cyclase (bPAC) activation in steroidogenic interrenal cells via blue light19,20. For the present study, we used the same
construct to generate a new transgenic line, _Tg(star:bPAC-2A-tdTomato)__uex300_, in a different strain, to obtain a higher cortisol level than previously feasible (Fig. 1a). After 10 min of
exposure to ambient white light (containing a blue light component), 5 days post fertilisation (dpf) _Tg(star:bPAC-2A-tdTomato)__uex300_ larvae exhibited elevated cortisol levels compared
to, their negative siblings which lack the transgene (Fig. 1b). Notably, _Tg(star:bPAC-2A-tdTomato)__uex300_ larvae shielded from blue light did not exhibit cortisol elevation, confirming
that bPAC activation by blue light is responsible for cortisol elevation (Fig. 1b). To assess the impact of elevated GC on brain development, we raised transgenic fish under standard
aquarium lighting, ensuring bPAC activation. This resulted in significantly elevated cortisol levels in _Tg(star:bPAC-2A-tdTomato)__uex300_ compared to wild types during larval stages (5 dpf
and 13 dpf, Fig. 1c)21. _Tg(star:bPAC-2A-tdTomato)__uex300_ raised under standard light conditions are denoted throughout as star:bPAC+ or bPAC + . To investigate the effects of elevated GC
on brain development we analyzed bulk RNA-sequencing (RNA-seq) data of whole brain samples from star:bPAC+ and wild-type fish at different developmental stages21. Gene Ontology (GO)
analysis of differentially expressed genes (DEGs) that were down-regulated in star:bPAC+ brains compared with wild types were enriched for a number of biological processes, including
Neurogenesis at all developmental stages (GO:0022008, adjusted _p_ value 2.47E-20 at 6 dpf, 5.45E-08 at 13 dpf, 1.11E-17 at 120 dpf). Up-regulated genes were not enriched for GO term
neurogenesis. To investigate the effects of GC exposure on neurogenesis during development, we focused downstream analysis on the early life time-points: 5/6 dpf and 13 dpf. 225 genes
associated with neurogenesis were down-regulated in star:bPAC+ brains at either 6 dpf or 13 dpf (Fig. 1d). To confirm our results and identify specific features of neurogenesis that are
altered by elevated GC, we performed qPCR on independent samples of whole brains from 5 dpf star:bPAC+ and wild types for genes associated with neurogenesis processes or distinct cell types.
Of the 14 categories of genes analysed, only one category was associated with consistent differential expression in star:bPAC+. Expression of _Pcna_, _mki67_ and _mcm5_, all associated with
cell proliferation, showed increased expression in star:bPAC+ compared with wild types (Supplementary Fig. 1). These data support that neurogenesis, and especially cell proliferation, is
altered by elevated GC. To identify whether the effect of GC on neurogenesis is developmentally dynamic, we performed unsupervised clustering analysis of neurogenesis DEGs at 6 dpf and 13
dpf and identified 6 different clusters. Cluster 2 and cluster 5 genes showed a relatively stronger down-regulation at 6 dpf compared to 13 dpf, meanwhile cluster 1 genes showed a relatively
stronger downregulation at 13 dpf (Fig. 1e). Cluster 3 genes were strongly downregulated at both time-points (Fig. 1e). Interestingly, cluster 4 and cluster 6 genes showed a time
point-specific down-regulation. Cluster 4 genes were downregulated in star:bPAC+ at 6 dpf but were not differentially expressed at 13 dpf (Fig. 1e). Meanwhile cluster 6 genes were
down-regulated in star:bPAC+ at 13 dpf but not at 6 dpf (Fig.1e). These results suggest that some effects of elevated GC on neurogenesis are temporarily dynamic during early life. GO
analysis of genes within the clusters indicated that all clusters contained genes associated with neuron differentiation (Fig. 1f). Gliogenesis and glial cell development were enriched in
multiple clusters. There were also some terms which were restricted to single or small numbers of clusters. For example, cluster 4 genes were associated with neuron fate commitment, cluster
3 with neuron migration, and cluster 2 with neuronal stem cell population maintenance (Fig. 1f). These results suggest that specific aspects of neurogenesis exhibit temporally dynamic
differential expression during early life as a result of GC exposure. PRECOCIOUS HYPOTHALAMIC DEVELOPMENT FOLLOWING ELEVATED GC To determine whether the effects of elevated GC on cell
proliferation were brain-wide or region specific we performed immunohistochemistry (IHC) for mitosis marker phospho-histoneH3 (pH3) on brains of 5 dpf star:bPAC+ and wild-type larvae (Fig.
2a). Cell counting of proliferating pH3+ cells across brain regions indicated a trend towards an increase in number of pH3+ cells in the valvula cerebelli (ce-v) of the hindbrain, however,
the only significant increase was restricted to the hypothalamus (Fig. 2b). We next sought to determine which cell types were associated with the increased proliferative capacity in the
hypothalamus of 5 dpf star:bPAC + . To this end, we performed Fluorescent in situ hybridisation (FISH) for _rx3_ (_retinal homeobox gene 3)_, combined with IHC for Pcna and pH3. _rx3_ is a
transcription factor known to be expressed in hypothalamic progenitor cells in zebrafish where it plays a critical role in hypothalamic development27, and its mammalian orthologue _Rax_ is
known to be expressed in hypothalamic radial glia28. Pcna and pH3 are markers for proliferating cells during S-phase and M-phase respectively. Pcna+ cells were situated around the 3rd
ventricle of the hypothalamus, especially around the lateral recess, in both star:bPAC+ and wild-type larvae, with a small number of pH3+ cells scattered throughout this region (Fig. 2c). We
distinguish the Pcna+ region around the hypothalamic ventricles as the ‘proliferative zone’ and observed co-localisation of _rx3_ with Pcna and pH3 within the proliferative zone (Fig. 2c,
white arrowheads). Quantitative analysis of confocal stacks revealed that the hypothalamus was significantly larger (Fig. 2f), and we found a significant increase in _rx3_ and Pcna signal
intensity, and significantly more pH3+ cells in the hypothalamus of 5 dpf star:bPAC+ compared with wild types (Fig. 2g–i). To determine whether the observed increase in hypothalamic
proliferation in the hypothalamus of 5 dpf star:bPAC+ larvae is a direct effect of developmental GC exposure, we treated star:bPAC+ larvae with either the GR antagonist RU-486 or MR
antagonist Spironolactone from 2 dpf until 5 dpf. Treatment with RU-486 did not significantly affect hypothalamic volume or hypothalamic Pcna signal, however it was sufficient to
significantly reduce the number of pH3+ mitotic cells in the hypothalamus (Supplementary Fig. 2). Meanwhile treatment of star:bPAC+ larvae with Spironolactone had no effect on hypothalamus
size, Pcna signal or pH3 cell count (Supplementary Fig. 2). This suggests that GR signalling is a significant driver of the increased hypothalamic proliferation observed in star:bPAC+
larvae. Further, we observed that the excess hypothalamic pH3+ cells were _rx3_ + , meanwhile _rx3_-/pH3+ cells were not different (Fig. 2i). These results suggest that excess proliferating
hypothalamic cells in star:bPAC+ are _rx3_-expressing radial glia. Interestingly, we identified two Glucocorticoid Response Elements (GREs) within the zebrafish _rx3_ gene (Fig. 2l). GRE1 is
in the 5’ UTR, which is most likely a regulatory region. ChIP-qPCR analysis revealed enrichment of GR antibody (compared with IgG) at both GREs in larval brain samples, supporting that GR
does indeed bind to regulatory regions of the _rx3_ gene (Fig. 2m). Further, enrichment of GR was higher in dexamethasone-treated wild-type samples, compared with wild-type controls, and
higher still in star:bPAC+ samples. We speculate that the increase in bPAC+ brains relative to dexamethasone-treated wild types is related to differences in dose, delivery, and rhythmicity
of exposure. This experiment shows that GC exposure directly regulates GR-mediated control of _rx3_ gene expression in the brain. Whilst _rx3_ appeared to be co-expressed with Pcna in
proliferating radial glia, we also analysed Blbp (brain lipid binding protein, fabp7a) localisation, which is known to be expressed in predominantly quiescent radial glia in the zebrafish
brain29. Consistent with this, we observed strong expression of Blbp in cells lining the third ventricle in the Pcna-negative rostral domains of the hypothalamus, whilst some Blbp signal was
also observed in Pcna-positive cells of the lateral recess (Fig. 2d). In 5 dpf star:bPAC+ larvae, we observed increased Blbp signal, in the basal hypothalamus (Fig. 2d, j), suggesting that
increased numbers of glial cells might include both proliferative and quiescent subtypes. Further, we observed increased signal for Otpa (orthopedia homeobox a) in the hypothalamus of 5 dpf
star:bPAC+ larvae (Fig. 2e, k), which is predominantly expressed in early post-mitotic neuronal precursors30, suggesting that the excess GC-exposed hypothalamic proliferative radial glia do
exit the cell cycle, leading to increased numbers of neuronal precursors. To assess the impact of excess hypothalamic progenitor cells and neuronal precursors at 5 dpf, we examined
differentiation into hypothalamic neurons. Hypothalamic expression of _agrp_ (_agouti-related peptide_), _avp_, _hcrt_, _pmch_ (_pro-melanin-concentrating hormone_), and _pomc_ showed no
difference between star:bPAC+ larvae and wild type (Supplementary Fig. 3). However, a decrease in hypothalamic _crhb_ (_corticotropin releasing hormone b)_ expression in star:bPAC+ larvae
was observed, consistent with heightened negative feedback to _crh_+ neurons due to elevated GC (Supplementary Fig. 3). Additionally, an increase in _npy_ + (_neuropeptide y_) cells in the
rostro-lateral intermediate hypothalamus, along with elevated cluster 3 and cluster 6 dopaminergic (DA, th + ) neurons in the hypothalamus/PT (posterior tuberculum), was noted in star:bPAC+
larvae compared to wild type (Supplementary Fig. 3). These findings suggest that the excess progenitor cells and neuronal precursors in star:bPAC+ larvae at 5 dpf may promote the
differentiation of specific neuronal subtypes in the developing hypothalamus. Together these data suggest that elevated GC drives precocious development of the hypothalamus in zebrafish
larvae. PROGENITOR CELLS FAIL TO DIFFERENTIATE IN THE HYPOTHALAMUS OF STAR:BPAC+ LARVAE Since we observed excess proliferating radial glia in the hypothalamus of 5 dpf star:bPAC+ larvae, we
sought to trace the proliferating cells of the hypothalamus after 5 dpf to determine their fate. To do this we exposed 4 dpf larvae to BrDU (Bromodeoxyuridine) overnight for 17 h until 5 dpf
and then fixed larvae at 5, 8, 10 or 13 dpf for double-IHC analysis of BrDU and Pcna (Fig. 3a). At 5 dpf we observed successful incorporation of BrDU into proliferating cells, as observed
by co-expression with Pcna, in the hypothalamus of both star:bPAC+ and wild-type larvae (Fig. 3b). By 8, 10 and 13 dpf, BrDU+ cells in the wild-type hypothalamus have begun exiting the cell
cycle and are now mostly observed around the perimeter of the Pcna-expressing proliferative zone (Fig. 3c, Supplementary Fig. 4). Indeed, we observed that the percentage of hypothalamic
BrDU+ cells that co-express Pcna drops from around 75% at 5 dpf to 50−55% at 8−13 dpf (Fig. 3d), suggesting that some of the BrDU+ cells that were proliferating at 5 dpf have lost their
progenitor cell status and differentiated. In stark contrast, hypothalamic BrDU+ cells in star:bPAC+ larvae are still predominantly observed within the Pcna+ proliferative zone after 5 dpf
(Fig. 3c, Supplementary Fig. 4). In fact, the percentage of hypothalamic BrDU+ cells that co-express Pcna remains at around 100% from 5 dpf until 13 dpf in star:bPAC+ larvae (Fig. 3d). This
indicates that in star:bPAC+ larvae, hypothalamic progenitor cells remain proliferative between 5 dpf and 13 dpf and do not differentiate. Further, we observed an increase in the number of
hypothalamic BrDU+ cells between 5 dpf and 8 dpf in wild types, supporting that these cells are dividing and therefore increase in number, however no such increase was observed in star:bPAC+
larvae (Fig. 3e). Similarly, the number of hypothalamic Pcna+ cells increased in wild types between 5 dpf and 10 dpf, indicating that the proliferative zone continues to expand as cells
divide during this period (Fig. 3f). No increase in the number of hypothalamic Pcna+ cells was observed in star:bPAC+ larvae during this developmental period (Fig. 3f), further supporting
that neurogenesis appears to stall during this developmental window in star:bPAC+ larvae. Together, these data support that in star:bPAC+ larvae, hypothalamic progenitor cells fail to
differentiate, and neurogenesis is stalled during the 5−13 dpf time window. FAILED HYPOTHALAMIC MATURATION FOLLOWING ELEVATED GC Since we observed that neurogenesis of proliferating
hypothalamic cells was altered between 5 dpf and 13 dpf, we investigated hypothalamic cell populations at 13 dpf. In the 13 dpf hypothalamus, _rx3_ was expressed in a subset of Pcna+ cells
around the lateral recess, and Blbp was expressed mainly along the midline 3rd ventricle anterior to the proliferative zone, in a similar manner to at 5 dpf (Fig. 4a). In 13 dpf star:bPAC+
larvae, we observed that the size of the hypothalamus was no longer larger than that of wild types, as we observed at 5 dpf (Fig. 4b). Expression of hypothalamic _rx3_ was low at this
developmental stage, and showed a trend towards a reduction in star:bPAC+ compared with wild type (Fig. 4a, c). In stark contrast to the 5 dpf hypothalamus, we now observed a dramatic
reduction in expression of Pcna in 13 dpf star:bPAC+ hypothalamus (Fig. 4a, d). Meanwhile, Blbp signal intensity was not different between 13 dpf wild-type and star:bPAC+ larvae (Fig. 4a,
e). Strikingly, when we performed co-expression analysis, we observed that whilst only a small proportion of Blbp+ cells was proliferative in 13 dpf wild-type hypothalamus (~10%), almost all
Blbp+ cells were quiescent in star:bPAC+ (Fig. 4f). Further, we observed that in wild types most hypothalamic _rx3+_ cells (~70%) were proliferative and expressed Pcna, however in
star:bPAC+ most _rx3+_ cells (~85%) were quiescent and didn’t express Pcna (Fig. 4g). These data support that proliferative radial glia are lost in the hypothalamus of 13 dpf star:bPAC+
larvae. Next, we sought to identify an explanation for the loss of proliferative radial glia in the hypothalamus. We first analysed cell death in the hypothalamus of 13 dpf larvae using a
TUNEL assay. We did not observe a significant difference in the number of apoptotic cells in the hypothalamus of 13 dpf star:bPAC+ larvae compared to wild types (Supplementary Fig. 5a),
suggesting that loss of hypothalamic radial glia is not primarily due to increased cell death. We speculate that the loss of proliferation in the star:bPAC+ hypothalamus might be due to
senescence of the formerly highly proliferative radial glia. In support of this hypothesis, we observed that one of the five significantly enriched KEGG pathways in our whole brain RNA-seq
data set in the 13 dpf star:bPAC+ was cellular senescence, a pathway which was not enriched in the 6 dpf star:bPAC+ samples (Supplementary Fig. 5b, c). In the RNA-seq analysis, 102 of 150
genes associated with the KEGG pathway cellular senescence were differentially expressed in 13 dpf star:bPAC+ brain samples, meanwhile at 6 dpf only 60 of those genes were differentially
expressed, and they generally showed a lesser degree of fold change (Supplementary Fig. 5d). This suggested that proliferative radial glia might have been lost in the hypothalamus of 13 dpf
star:bPAC+ larvae due to cellular senescence, however further investigation of senescent markers in the hypothalamus is required to confirm this. Additionally, we confirmed that the loss of
hypothalamic proliferation is maintained into later life, since we observed that in juvenile star:bPAC+ fish, the hypothalamus is smaller and contained fewer mitotic cells (Supplementary
Fig. 6a–c). This suggests that the proliferative hypothalamic cells lost at 13 dpf do not re-enter the cell cycle later in development, suggesting that they have become senescent, rather
than quiescent. Since we observed a dramatic reduction in cell proliferation in the hypothalamus of 13 dpf star:bPAC+ larvae, we postulated that downstream neurogenesis and neuronal
differentiation might also be affected. Firstly, analysis of Otpa+ neural precursor cells at 13 dpf revealed no difference between star:bPAC+ and wild types (Supplementary Fig. 7a, b). We
next analysed hypothalamic neuronal subtypes in 13 dpf star:bPAC+ and wild type brains. _agrp_+ cells in the arcuate nucleus were significantly reduced in star:bPAC+ larvae (Fig. 4h, i).
Similarly, _cart4_ + (_cocaine- and amphetamine-regulated transcript-4_) cells in the rostro-lateral intermediate hypothalamus showed a decrease in star:bPAC+ larvae (Fig. 4h, j). Although a
trend of reduction was observed in _hcrt_+ cells in the dorso-rostral hypothalamus of star:bPAC+ larvae (Fig. 4h, k), no differences were noted in _npy_ + , _pomca_ + , or _th_+ cells
(Supplementary Fig. 7a, c, e, f). Notably, a significant increase in _pmch_+ cells was observed in the rostro-lateral intermediate hypothalamus of star:bPAC+ larvae compared to wild type
(Supplementary Fig. 7a, d). These findings suggest that prolonged exposure to GC until 13 dpf affects neuronal differentiation and potentially disrupts the balance of hypothalamic neuronal
subtypes in star:bPAC+ larvae. FUNCTIONAL CONSEQUENCES OF ELEVATED GC: PRECOCIOUS FEEDING FOLLOWED BY PHYSICAL DECLINE The increased size and proliferative capacity of the hypothalamus in 5
dpf star:bPAC+ larvae suggested that development of the hypothalamus is accelerated in these animals. Therefore, we hypothesised that behaviours associated with the hypothalamus that develop
around this developmental stage might also be altered and thus we analysed feeding in 5 dpf larvae. Feeding usually emerges at around 120 hpf (5 dpf) in lab-raised zebrafish larvae, and
larvae are capable of eating small live food. We exposed 5 dpf larvae to live rotifers and saw that star:bPAC+ larvae consumed significantly more rotifers during the trial (Fig. 5a,
Supplementary video 1), suggesting that GC-induced precocious hypothalamic development is accompanied by early emergence of an associated behaviour, feeding. In stark contrast to 5 dpf, at
13 dpf star:bPAC+ larvae fed less than wild types (Fig. 5b). Interestingly, in 13 dpf wild-type larvae we observed that food intake positively correlated with body size, in that larger
larvae ate more, whilst in star:bPAC+ larvae no significant trend was apparent. (Fig. 5c). Hence, star:bPAC+ larvae consumed less, regardless of their size. Whilst at 5 dpf and 13 dpf we did
not observe any difference in larval body size, at 28 dpf and 120 dpf star:bPAC+ fish were significantly smaller than wild types (Fig. 5d). This suggests that the reduction in food intake
in star:bPAC+ animals from 13 dpf ultimately leads to impaired growth. Finally, we also observed reduced fertilisation rates and reduced long-term survival of star:bPAC+ fish (Fig. 5e, f).
Together, these results point towards potential functional consequences of failed hypothalamic development after 5 dpf, including impaired behaviour and an early physical decline. Further
work is required to directly link altered hypothalamic neurogenesis with the observed functional alterations in GC-exposed animals. DISCUSSION This work shows that elevated glucocorticoid
alters the trajectory of hypothalamic development and function. We identify that the hypothalamus is a highly GC-sensitive region where elevated GC causes precocious development followed by
failed maturation and early decline accompanied by impaired feeding, growth, and survival. Our data supports that, at least in some cases, reduced cell proliferation following GC exposure
might be related to a prior increase in proliferation. This aligns with the concept of GC-inducible stem cells31,32. Previous studies have demonstrated GC-induced proliferation in various
contexts33,34,35,36, resembling mechanisms observed during brain injury, where quiescent radial glia enter the cell cycle in response to trauma29,37. Few studies have explored the
developmental impact of GC on cell proliferation. Noorlander et al. observed that antenatal GC treatment in mice led to an initial reduction in embryonic hippocampal proliferation, followed
by increased proliferation postnatally and subsequent reduction in adulthood38. Similarly, mouse models of ELS exhibited an initial increase in hippocampal proliferation followed by a later
decline10,12. A meta-analysis on age-dependent effects of ELS indicated a negative correlation between age and changes in proliferation, suggesting that ELS-exposed animals exhibit increased
proliferation early in development, followed by a reduction in later life39. It is possible that our interpretation of the results presented here also applies to other studies, in that GC
drives precocious proliferation leading to failed maturation and a rapid decline. In our model, we observed profound effects on a population of Neural stem/progenitor cells (NSPCs) within
the proliferative zone of the developing hypothalamus. This population of hypothalamic radial glia might be akin to tanycyte cells observed in mammals, which also express _Rax_/_rx3_ and
_Blbp_18, and were recently shown to be sensitive to ELS exposure in adult mice17. We postulate that these cells might be especially sensitive to elevated GC, since they are adjacent to the
3rd ventricle, potentially detecting elevated GC levels within the CSF. The increased sensitivity of the hypothalamus to GC could also be influenced by higher levels of GR, however, it
remains unclear whether the zebrafish hypothalamus expresses higher GR levels than other brain regions. The specificity of the proliferation phenotype observed in our study lacks a clear
explanation. While it’s uncertain if the hypothalamus is uniquely affected in our model, it’s plausible that GC effects on different brain regions may emerge over different time scales.
Previous research injecting cortisol into zebrafish at the 1-cell stage, mimicking maternal cortisol transfer, demonstrated increased neurogenesis in specific brain regions, such as the
pallium and preoptic region, but not in the rostral hypothalamus of larvae at 5 dpf40. This supports that GC can induce neurogenesis in specific brain regions, likely depending on the
context of the exposure. Developmental GC exposure in star:bPAC+ larvae significantly affected feeding behaviour, with increased consumption at 5 dpf and decreased consumption at 13 dpf,
persisting into adulthood21. These changes correlated with an early increase in proliferative hypothalamic radial glia at 5 dpf, diminishing by 13 dpf, accompanied by a reduction in
feeding-regulating neurons expressing _agrp_, _cart_, and _hcrt_ at 13 dpf. In larval zebrafish, hypothalamic _agrp_ neurons are known to stimulate feeding41,42, and orexin, produced by
_hcrt_ neurons, is also known to stimulate food intake in zebrafish43. Meanwhile the effects of _cart_ on appetite are more complex44. We also observed an increase in _pmch_+ neurons at 13
dpf. Whilst the role of MCH is not fully understood in zebrafish, its expression is known to increase in response to fasting in zebrafish45, and in goldfish it is anorexigenic46. As such, it
is likely that the observed reduction in numbers of _agrp_ and _hcrt_ neurons and increase in _pmch_ neurons in 13 dpf star:bPAC+ larvae contribute to the observed reduction in food
consumption at the same developmental stage. At 5 dpf, increased feeding in star:bPAC+ larvae coincided with reduced _crh_+ neurons and an increase in _npy_+ neurons. Although CRH neurons
can stimulate food intake47, evidence indicates that in the arcuate nucleus CRH may inhibit appetite-stimulating _agrp_ + /_npy_+ neurons, thus stimulating food intake48. Thus, in our model,
it is plausible that reduced _crh_ expression might dis-inhibit _npy_ expression, leading to the observed increase in _npy_+ neurons, subsequently stimulating food intake in 5 dpf star:bPAC
+ . Previous work also supports that hypothalamic _th_+ neurons stimulate food intake by regulating activity of _pomc_+ and _npy_ + /agrp+ neurons49, suggesting that the increased _th_ in
specific hypothalamic DA clusters may also play a role in the early increase in food consumption we observed. Another hypothesis regarding the basis of the altered food consumption in the
star:bPAC+ larvae relates to the observed changes in hypothalamic radial glia. The population of hypothalamic radial glia altered by developmental GC exposure in our model display
similarities to mammalian tanycyte cells, based on location, morphology, and marker expression18. Recent work demonstrated that hypothalamic tanycytes can regulate food intake50.
Hypothalamic tanycytes are known to project to neighbouring orexigenic _npy_/_agrp_ and anorexigenic _pomc_ neurons of the arcuate nucleus, and tanycyte activation was shown to depolarise
these neurons in vitro, and lead to hyperphagia in vivo. It is interesting to postulate that if similar functionality occurs in the hypothalamic radial glia of zebrafish, then the observed
increase in these cells at 5 dpf might contribute to the increased feeding behaviour observed, meanwhile the subsequent reduction of these cells might contribute to the reduction of feeding
behaviour observed in 13 dpf star:bPAC+ larvae. Further work is required to characterise the development and function of the zebrafish hypothalamic radial glia, and to determine the
mechanisms underlying the altered feeding observed in star:bPAC+ larvae. Whilst the reduced food intake in 13 dpf star:bPAC+ larvae potentially contributes to the subsequent decline in
growth, activation of GR signalling is known to supress growth via increased protein catabolism, and this is potentially also a contributing factor in our model51. We hypothesise that the
precocious hypothalamic development observed here may represent an example of GC-induced adaptive plasticity (Fig. 6), as observed in other animals. For example in red squirrels, females
exposed to high-density cues (mimicking increased competition) increase their levels of endogenous GC which drives them to produce offspring that grow faster than controls52. This so-called
adaptive plasticity is acknowledged to have short-term fitness benefits, but the investment is often associated with costs in later life53. The costs of accelerated development can include
reduced lifespan, which is known as the growth-rate lifespan trade-off54, as well as reductions in breeding success55. Interestingly, there is evidence directly linking divergence from the
normal growth trajectory with lifespan, since sticklebacks that are forced to grow faster have a reduced lifespan, whilst those in which growth was slowed down lived longer than controls54.
In our model, chronic GC exposure ultimately appears to lead to allostatic overload, a maladaptive state whereby an organism can no longer adapt to its conditions56. Whilst GC-induced
acceleration of growth-rate leading to adverse phenotypes later is widely observed at the organism level, the underlying cellular and molecular mechanisms are poorly understood. Our work
provides new insight into these processes in the developing brain. However, the potential evolutionary benefit of the precocious hypothalamic development and early feeding observed in our
study requires further investigation. In the context of the early-late life trade-off, the subsequent reduction in breeding success and/or lifespan is thought to be related to oxidative
stress57,58. Exposure to elevated GC is known to increase oxidative stress59, a phenomenon in which an over-accumulation of reactive oxygen species (ROS) occurs in cells and tissues60. ROS
is known to cause cellular damage, especially to DNA, and can lead to accelerated telomere shortening61. In a previous study of amphibian larvae, the relationship between stress,
development, growth and aging depended on the type of stressor exposure62. Predator exposure initiated faster development and enhanced growth of survivors, but they showed signs of oxidative
stress, had shorter telomeres and reduced long-term survival62. This indicates that stress exposure may initiate faster development in early life, but at a cost to long-term health and
lifespan. Telomeres shorten with every round of cell division and when a critical size is reached this imposes a functional limit on cell replication, leading to replicative senescence,
which can contribute to age-related diseases61. One possibility in our model is that the excess proliferation of hypothalamic progenitors observed in 5 dpf star:bPAC+ larvae leads to
subsequent senescence of those cells. In contrast to quiescent, apoptotic, or terminally differentiated cells, senescent cells have incurred an irreversible cell cycle arrest, yet remain
viable, have alterations in metabolic activity and undergo dramatic changes in gene expression63. In support of our hypothesis, a previous study of liver progenitor cells reported that GC
exposure induced cell proliferation leading to long-term replicative senescence/stemness exhaustion64, and GC is also reported to induce senescence of NSPCs in vitro65. The presence of
senescent cells in the proliferative zone of the GC-exposed hypothalamus at 13 dpf might explain why we observed reduced expression of Pcna and _rx3_, and a reduced proliferative capacity of
the remaining glial cells in the proliferative zone of star:bPAC+ larvae. Telomere shortening and senescence are associated with aging and age-related diseases; and models of accelerated
aging present with phenotypes such as reduced survival and physical decline66, as observed in our model. Additionally, chronic exposure to GC is associated with signs of accelerated aging at
a cellular level67. Further analysis of oxidative stress and telomere homoeostasis would provide further insight into the mechanisms underlying the phenotypes observed here. We conclude
that the phenotypes observed in our optogenetic model are the result of exposure to elevated GC produced in the interrenal, rather than other interrenal steroid hormones, which are not
detectable in larval zebrafish21,23,68. We propose that GC-induced alteration of hypothalamic development observed in our model is mediated by GR-regulated gene transcription. In support of
this, we showed that treatment with GR antagonist RU-486 reduced pH3+ mitotic cells in the star:bPAC+ hypothalamus. Whilst treatment with spironolactone had no effect on proliferation,
whether spironolactone acts as an MR antagonist in fish, as it does in humans, is unclear69. GR expression in the brain is relatively ubiquitous70, however, in the mouse, MR expression is
mostly restricted to neurons and some astrocytes of the hippocampus and is not expressed in NSPCs71,72, supporting that MR is unlikely to mediate the altered cell proliferation observed in
our model. Conversely, MR is known to play a role in differentiation. MR overexpression stimulates neuronal differentiation71,73 and MR is known to play a role in hippocampal neuron fate and
associated behaviours74. As such, it is possible that MR mediates the altered hypothalamic neuron phenotypes and behavioural changes observed in our model, however, this requires further
investigation. We also provide evidence for regulation of hypothalamic _rx3_ by GR. Indeed, GR signalling is known to regulate cell cycle progression in NSPCs by inducing a specific pattern
of DNA methylation during aging75. In a model of replicative senescence, GC exposure lead to altered methylation of GR target gene _fkbp5_, which was exacerbated by age, and the subsequently
increased _fkbp5_ expression was associated with inflammation and myocardial infarction in humans, suggesting that GC is linked with age-related disease via a mechanism involving epigenetic
regulation of _fkbp5_76. Meanwhile in cancer cells, GC exposure can induce a cell dormancy state which is in part senescence and in part quiescence, mediated by GR target gene _CDNK1C_
(_cyclin-dependent kinase inhibitor 1C)_77, further supporting that GC exposure might regulate cell proliferation in our model via GR-mediated regulation of cell cycle. We speculate that the
effects of elevated GC on the hypothalamic developmental trajectory reported here might be an example of allostatic overload. Allostasis is the process of activating adaptive mechanisms in
response to external or internal changes, however prolonged exposure to stressors can lead to accumulation of so-called allostatic load, and subsequently a maladaptive state of allostatic
overload8, which is associated with age-related diseases and reduced lifespan. In support of this, it has been proposed that effects of stress or GC on NSPCs in young individuals may affect
their renewal potential in the long-term, predisposing to adult disease31. We hypothesise that whilst precocious hypothalamic development induced by GC might be an example of adaptive
plasticity in the short-term, the cost of this may be paid in later life through subsequent replication senescence of those cells, leading to failed hypothalamic maturation, subsequent
impaired feeding behaviour and ultimately physical decline (Fig. 6). Further understanding of how stress and GC exposure can alter developmental trajectories at the molecular and cellular
level is of critical importance to reduce the burden of mental and physical ill health across the life-course. METHODS ZEBRAFISH HUSBANDRY AND MAINTENANCE Zebrafish experiments were
performed at the University of Exeter Aquatic Resource Centre and at the Johannes Gutenberg University of Mainz in compliance with local and national animal welfare laws, guidelines, and
policies and approved by local government (Landesuntersuchugsamt Rheinland-Pfalz, Germany – 23 177-07/G20-1-033 and UK Home Office PPL number: PEF291C4D).
_Tg(2kbStAR:bPAC-2A-tdTomato)__uex300_ were bred with wild-type TU zebrafish strain and were screened at 4 dpf on a fluorescent stereomicroscope. Adult zebrafish were maintained under
standard zebrafish husbandry conditions, under a 12:12 light/dark cycle78. Transgenic fish exhibited normal sex ratios and displayed no gross developmental abnormalities. Until 5 dpf larvae
were maintained at 50 larvae per Petri dish in 40 ml system water, after which they were maintained until 13 dpf in 200 ml system water. For most experiments, larvae at 5 dpf, 8 dpf, 13 dpf,
28 dpf were used and sex is not yet determined. For fertilisation rate only males were used. For body size analysis at 120 dpf, equal numbers of males and females were used. RNA SEQUENCING
RNA sequencing data was obtained from our other study21, which is deposited in the European Nucleotide Archive (ENA, PRJEB53713). The run accession IDs for the RNA-seq data reported in this
study are: TU WT 6 dpf: ERR10476787 - ERR104767 91, _star_:bPAC positive 6 dpf: ERR10476 807 - ERR10476 811; TU WT 13 dpf: ERR104767 92 - ERR104767 96; _star_:bPAC positive 13 dpf: ERR10476
812 - ERR10476 816. The detailed protocol for sample collection, RNA preparation, mRNA sequencing, and bioinformatic analysis is available in an online methods repository79. GENE ONTOLOGY
ANALYSIS The functional enrichment analysis was performed using g:Profiler (version _e107_eg54_p17_bf42210)_ with an adjusted _p_ value of 0.05. For GO analysis of neurogenesis gene clusters
(Fig. 1e), significant GO terms for each cluster were manually slimmed according to QuickGO ancestor charts. Heatmaps were generated in R using the heatmap.2 function. QPCR Fish were
immobilised using ice-cold water and larvae or dissected juvenile/adult brains were stored in RNA later solution. Sample collection was carried out between 08:00 and 10:00. Larvae were
subsequently dissected, such that each replicate consisted of 15 larval heads (eyes and jaw removed) or 3 juvenile/adult brains. RNA was extracted using TRIzol™ Reagent (Invitrogen™,
15596026), as previously described80, and cDNA was synthesised using High-Capacity RNA-to-cDNA™ Kit (Applied Biosystems, 4387406). Approximately 100 ng cDNA was used in a 10 μl qPCR reaction
with PowerUp™ SYBR™ Green Master Mix (A25778, Applied Biosystems). Primer sequences can be found in Supplementary Data 2. The reactions were run in Hard-Shell 96-Well PCR Plates (HSP9601,
BioRad) on a CFX96 Real Time PCR machine (Bio-Rad) using a standard protocol. Relative expression was calculated using the 2–∆∆Ct method, with _18_ _s_ or _sep15_ as a reference gene.
FLUORESCENT IN SITU HYBRIDISATION (FISH) AND IMMUNOHISTOCHEMISTRY (IHC) The FISH and IHC methods were based on a protocol by Jakob von Trotha (2016) for whole zebrafish embryos and larvae.
Larvae or juvenile fish were immobilised in ice-cold water and then whole larvae or dissected juvenile fish brains were fixed in 4% PFA overnight at 4 °C, dehydrated in methanol and
subsequently stored at −20 °C. After rehydration, larvae or brains were permeabilised using proteinase K (10 μg/ml) treatment at 37 °C and then re-fixed in PFA. DIG-conjugated mRNA probes
were hybridised overnight at 65 °C. In situ hybridisation probes for _rx3_, _agrp_, _cart4, npy, pmch, th, crhb, avp_ were synthesised using primers listed in Supplementary Data 2. _pomc_
probe was synthesised from a plasmid, as previously described81. After removal of probe and washing, samples were blocked for 2 h in blocking solution (1% blocking reagent; (Roche,
11096176001) in malic acid buffer (0.15 M maleic acid, 0.15 M NaCl, PH 7.5)) and then incubated overnight at 4 °C in anti-DIG POD antibody (Roche, 11207733910) diluted 1:300 in blocking
solution. After more washing, samples were incubated for 40 min in the dark with 1:200 FITC-tyramide (synthesised from product 46410, Thermo Scientific), 0.003% H2O2, 2% dextran sulphate in
PBST. For IHC antigen retrieval was performed according to published methods82. Samples were blocked in 10% Normal goat serum in PBST for 2 h before incubation in primary antibody overnight
at 4 °C. For IHC of BrDU-treated samples, samples were treated with 1 N HCl for 30 min prior to blocking. Primary antibodies used were: pH3 (Merck/Millipore, 06-570), Pcna (Sigma, MABE288),
BLBP (Sigma, ABN14), Otpa83, BrDU (abcam, ab6326). After washing all day, samples were incubated overnight in 1:1000 secondary antibody with 1:200 hoechst (H3570, Invitrogen). Secondary
antibodies used were Goat anti-Rabbit IgG (H + L) Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 488 (A-11008, Invitrogen), Goat anti-Rabbit IgG (H + L) Cross-Adsorbed Secondary Antibody,
Alexa Fluor 633 (A-21070, Invitrogen), Goat anti-Mouse IgG (H + L) Cross-Adsorbed Secondary Antibody, Alexa Fluor 568 (A-11004, Invitrogen), Goat anti-Rat IgG (H + L) Cross-Adsorbed
Secondary Antibody, Alexa Fluor 488 (A-11006, Invitrogen). After staining all samples were washed further then cleared overnight in glycerol. Samples were imaged on a Zeiss LSM 880 confocal
microscope using a 25x or 10x objective. IMAGE PROCESSING Image processing was performed in Fiji (ImageJ2, version 2.9.0). For whole brain counting of cells, z-stacks were taken across the
brain from dorsal to ventral and regions were defined using the hoechst staining with the Z-Brain viewer84. Images were thresholded and pH3 cells in each region were manually counted in a
blinded manner. For counting of cells within the hypothalamus, cell counts were performed manually from z-stacks, in a blinded manner. Similarly, hypothalamic volume was calculated based on
hoechst staining in z-stacks, using Z-Brain viewer. Corrected Total Cell Fluorescence (CTCF) was calculated as Integrated Density of cells – (Area of cells X Mean fluorescence of background
reading). For CTCF calculations, labelled cells were identified first by thresholding each image and creating ROIs of the labelled cells using the Analyze Particles function. The total
Integrated density and total area of all labelled cells was then used to calculate CTCF for each sample. For co-expression analysis, ROIs of labelled cells were generated for each marker as
described above. Percentage overlap was calculated using the Analyze Particles function with ROIs of marker 1 cells against a mask of marker 2 cells. Total area of cells was calculated from
the thresholded mask/ROIs of labelled cells. pH3 cells were manually scored as _rx3_- or _rx3_+ using the ROIs/masks as described above, in a blinded manner. All measures are normalised
relative to size of the hypothalamus. For the BrDU experiment, BrDU+ and Pcna+ cells are reported as the area of labelled cell coverage in μM. This was calculated from the thresholded
fluorescent signal for each marker within the hypothalamus. FEEDING BEHAVIOUR Prior to behavioural analysis, larvae were transferred to 24 well cell culture plates (83.3922, Sarstedt) with 1
larva per well in 1.5 ml aquarium water. Larvae were then left to acclimate to the behavioural testing room (maintained at 28 °C) for 2 h. 5 dpf larvae were fed 1 ml of diluted live
rotifers (approximately 20), meanwhile 13 dpf larvae were fed 5 live artemia. Larvae were recorded for 10 min using Basler Video Recording Software. The camera used was a Basler (Germany)
acA1300-200um USB 3.0 camera with ON Semiconductor PYTHON 1300 CMOS sensor (203 frames per second at 1.3 MP resolution) with a Computar Zoom Lens 18-108/2.5 (Japan). Number of successful and
unsuccessful prey captures and latency to first hunting attempt were analysed manually from video recordings in a blinded manner. BRDU EXPERIMENT Larvae were transferred to a 12 well dish
with 10 larvae per well. Larvae were incubated overnight from 4 dpf in 3 ml 10 mM 5-Bromo-2′-deoxyuridine (B5002, Sigma) with 1% DMSO for 17 h. On the morning of 5 dpf, BrDU was removed by
washing 3-times with aquarium water. Larvae were subsequently fixed in PFA or raised until a later stage for fixation. DRUG TREATMENTS Larvae were incubated from 6 hpf until 120 hpf with 50
μM dexamethasone (Sigma-Aldrich D2915) or from 48 hpf until 120 hpf with 2 μM RU-486 (Sigma-Aldrich, M8046) or with 10 μM Spironolactone (Sigma-Aldrich S3378). DMSO solution was used as a
control with 0.008% DMSO for Spironolactone control and 0.004% DMSO for Mifepristone control. Solutions were changed daily. Dexamethasone concentration was determined based on a previous
study that found that 50 μM dexamethasone could reliably induce negative feedback to the HPA axis85. RU-486 concentration was selected based on a previous study of GR mutant zebrafish86. The
spironolactone concentration used was selected based on a previous study87. CHIP-QPCR We analysed the sequence for the _rx3_ gene (ENSDARG00000052893,
chromosome:GRCz11:21:10755554:10759823:1). Primers for _rx3_ GRE1 and GRE2 are described in supplementary data 2 and were designed using Primer3. GR bounded chromatin was prepared following
the protocol from Idilli et al.88, with minor modifications. For each sample, 100 dissected 5 dpf larval heads (without eyes) were incubated for 10 min at room temperature in 1% formaldehyde
in PBS with protease inhibitors (A32955, Thermo Scientific) for cross-linking. We used a glucocorticoid receptor polyclonal antibody (24050-1-AP, Proteintech) and Rabbit IgG Isotype Control
(10500 C, Invitrogen) as primary antibodies. For each sample, 3 technical replicates were run on the PCR machine. TUNEL ASSAY Apoptotic cells were labelled using the In Situ Cell Death
Detection Kit, Fluorescein (11684795910, Roche). Larvae were first fixed, dehydrated, rehydrated and permeabilised as per the FISH protocol. Larvae were incubated in TUNEL reaction mix at 37
°C for 2 h and subsequently, washed and cleared as per the FISH protocol. FERTILISATION ANALYSIS Fertilisation rate of adult star:bPAC+ males was determined during standard pair matings
with wild type females and compared with wild type crosses. The fish used were 11 months of age. Embryos were collected from 19 wild type and 16 bPAC+ pairs across 3 independent mating
trials and successful fertilisation was calculated at 6−8 hpf by identifying the number of unfertilised embryos in each Petri dish. The fertilisation rate for each pair was determined by
averaging across all Petri dishes of embryos that came from each pair. Survival of fertilised embryos was confirmed at 24 hpf. SURVIVAL ANALYSIS Survival of star:bPAC+ and wild type fish was
determined by calculating the number of fish that survived in a stock from when larvae were placed in the facility nursery at 5 dpf until 2 months of age when fish were prepared for
transfer to the main aquarium facility. Survival was calculated from 9 wild type and 8 bPAC+ stocks that were placed into the nursery, where each stock consisted of between 40 and 300 fish
that were housed across multiple tanks at the same stocking density. These data were collected from different stocks that were raised over a duration of more than 2 years. CORTISOL A
step-wise method is reported in an online repository79. On the day prior to cortisol analysis, groups of 12 larvae were pipetted into each well of a 6-well plate with 6 ml system water per
well. For filtered light treatments the plate was stored inside a custom-built box covered by 550 nm long-pass filters (Thor- labs) and sampling was performed under filtered light. For acute
light exposure of filter-raised larvae, the lid was removed from the filter box whilst the box is inside the lit incubator, and larvae were exposed to white light for 10 min. Whilst still
inside the incubator, larvae were immobilised using ice water, transferred to an Eppendorf, and the water was removed. Samples were snap frozen on liquid nitrogen. The cortisol assay (Cisbio
HTRF Cortisol Kit, 62CRTPEG) was performed following the manufacturer’s protocol. Signal was detected using a CLARIO star plate reader (BMG Labtech). STATISTICS AND REPRODUCIBILITY All
analyses and graphs were generated in R (RStudio 2022.07.2). Prior to testing for statistically significant differences between groups, data were tested for normality and variance. Where
data did not fit the assumptions for t-testing or ANOVA, non-parametric alternatives were used. Where appropriate, Bonferroni correction for multiple testing was used. In all bar graphs mean
± standard error is presented for each group. Sample sizes consisted of between 4 and 23 biological replicates per group for microscopy experiments. For the RNA sequencing experiment, there
were 5 pools of brain samples per group. For cortisol experiments there were between 5 and 9 pools of larvae per group. For ChIP-qPCR there was 1 pool of biological replicates per group,
run as 3 technical replicates. For functional studies there were between 8 and 19 replicates per group. Exact sample sizes and number of replicates are indicated in each figure legend.
REPORTING SUMMARY Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY All sequenced reads for RNA-seq were
deposited in European Nucleotide Archive as part of our other study21 (ENA, PRJEB53713). The run accession IDs for the RNA-seq data reported in this study are: TU WT 6 dpf: ERR10476787 -
ERR104767 91, _star_:bPAC positive 6 dpf: ERR10476 807 - ERR10476 811; TU WT 13 dpf: ERR104767 92 - ERR104767 96; _star_:bPAC positive 13 dpf: ERR10476 812 - ERR10476 816. Source data for
figures are available in Supplementary data file 1. All other data are available from the corresponding author on reasonable request. CODE AVAILABILITY The code used for bioinformatic
analysis of the RNA sequencing data is available in an online methods repository79. REFERENCES * Chrousos, G. P. Stress and disorders of the stress system. _Nat. Rev. Endocrinol._ 5, 374–381
(2009). Article CAS PubMed Google Scholar * McKinlay, C. J. D., Dalziel, S. R. & Harding, J. E. Antenatal glucocorticoids: where are we after forty years? _J. Dev. Orig. Health
Dis._ 6, 127–142 (2015). Article CAS PubMed Google Scholar * Wolford, E. et al. Associations of antenatal glucocorticoid exposure with mental health in children. _Psychol. Med._ 50,
247–257 (2020). Article PubMed Google Scholar * Ilg, L., Klados, M., Alexander, N., Kirschbaum, C. & Li, S.-C. Long-term impacts of prenatal synthetic glucocorticoids exposure on
functional brain correlates of cognitive monitoring in adolescence. _Sci. Rep._ 8, 7715 (2018). Article PubMed PubMed Central Google Scholar * Davis, E. P., Sandman, C. A., Buss, C.,
Wing, D. A. & Head, K. Fetal glucocorticoid exposure is associated with preadolescent brain development. _Biol. Psychiatry_ 74, 647–655 (2013). Article CAS PubMed PubMed Central
Google Scholar * Bale, T. L. et al. Early life programming and neurodevelopmental disorders. _Biol. Psychiatry_ 68, 314–319 (2010). Article PubMed PubMed Central Google Scholar *
Lupien, S. J., McEwen, B. S., Gunnar, M. R. & Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. _Nat. Rev. Neurosci._ 10, 434–445 (2009). Article
CAS PubMed Google Scholar * Danese, A. & McEwen, B. S. Adverse childhood experiences, allostasis, allostatic load, and age-related disease. _Physiol. Behav._ 106, 29–39 (2012).
Article CAS PubMed Google Scholar * Moisiadis, V. G. & Matthews, S. G. Glucocorticoids and fetal programming part 1: outcomes. _Nat. Rev. Endocrinol._ 10, 391–402 (2014). Article
CAS PubMed Google Scholar * Suri, D. et al. Early stress evokes age-dependent biphasic changes in hippocampal neurogenesis, BDNF expression, and cognition. _Biol. Psychiatry_ 73, 658–666
(2013). Article CAS PubMed Google Scholar * Mirescu, C., Peters, J. D. & Gould, E. Early life experience alters response of adult neurogenesis to stress. _Nat. Neurosci._ 7, 841–846
(2004). Article CAS PubMed Google Scholar * Naninck, E. F. et al. Chronic early life stress alters developmental and adult neurogenesis and impairs cognitive function in mice.
_Hippocampus_ 25, 309–328 (2015). Article CAS PubMed Google Scholar * Lemaire, V., Koehl, M., Le Moal, M. & Abrous, D. Prenatal stress produces learning deficits associated with an
inhibition of neurogenesis in the hippocampus. _Proc. Natl. Acad. Sci._ 97, 11032–11037 (2000). Article CAS PubMed PubMed Central Google Scholar * Sawamoto, A. et al.
3,5,6,7,8,3′,4′-Heptamethoxyflavone, a citrus flavonoid, ameliorates corticosterone-induced depression-like behavior and restores brain-derived neurotrophic factor expression, neurogenesis,
and neuroplasticity in the hippocampus. _Molecules_ 21, 541 (2016). Article PubMed PubMed Central Google Scholar * Lehmann, M. L., Brachman, R. A., Martinowich, K., Schloesser, R. J.
& Herkenham, M. Glucocorticoids orchestrate divergent effects on mood through adult neurogenesis. _J. Neurosci._ 33, 2961–2972 (2013). Article CAS PubMed PubMed Central Google
Scholar * Youssef, M. et al. Early life stress delays hippocampal development and diminishes the adult stem cell pool in mice. _Sci. Rep._ 9, 4120 (2019). Article PubMed PubMed Central
Google Scholar * Bielefeld, P. et al. Early life stress decreases cell proliferation and the number of putative adult neural stem cells in the adult hypothalamus. _Stress_ 24, 189–195
(2021). Article CAS PubMed Google Scholar * Goodman, T. & Hajihosseini, M. K. Hypothalamic tanycytes-masters and servants of metabolic, neuroendocrine, and neurogenic functions.
_Front. Neurosci._ 9, 387 (2015). Article PubMed PubMed Central Google Scholar * Gutierrez-Triana, J. A. et al. Manipulation of interrenal cell function in developing zebrafish using
genetically targeted ablation and an optogenetic tool. _Endocrinology_ 156, 3394–3401 (2015). Article CAS PubMed Google Scholar * Nagpal, J., Eachus, H., Lityagina, O. & Ryu, S.
Optogenetic induction of chronic glucocorticoid exposure in early-life impairs stress-response in larval zebrafish. _bioRxiv_, 2022.2009.2009.507267 (2022). * Choi, M-K. et al. Lifelong
molecular consequences of high Glucocorticoids exposure during development. _bioRxiv_, 2023.2002.2013.528363 (2023). * Santulli, G. _Adrenal Glands: From Pathophysiology to Clinical
Evidence_ (Nova Biomedical, 2015). * Eachus, H. et al. Genetic disruption of 21-Hydroxylase in Zebrafish causes interrenal hyperplasia. _Endocrinology_ 158, 4165–4173 (2017). Article CAS
PubMed PubMed Central Google Scholar * Oakes, J. A. et al. Ferredoxin 1b deficiency leads to testis disorganization, impaired spermatogenesis, and feminization in Zebrafish.
_Endocrinology_ 160, 2401–2416 (2019). Article CAS PubMed Google Scholar * Li, N., Oakes, J. A., Storbeck, K.-H., Cunliffe, V. T. & Krone, N. P. The P450 side-chain cleavage enzyme
Cyp11a2 facilitates steroidogenesis in zebrafish. _J. Endocrinol._ 244, 309–321 (2020). Article CAS PubMed Google Scholar * Oakes, J. A., Barnard, L., Storbeck, K.-H., Cunliffe, V. T.
& Krone, N. P. 11β-Hydroxylase loss disrupts steroidogenesis and reproductive function in zebrafish. _J. Endocrinol._ 247, 197–212 (2020). Article CAS PubMed Google Scholar * Muthu,
V., Eachus, H., Ellis, P., Brown, S. & Placzek, M. Rx3 and Shh direct anisotropic growth and specification in the zebrafish tuberal/anterior hypothalamus. _Development_ 143, 2651–2663
(2016). CAS PubMed PubMed Central Google Scholar * Pak, T., Yoo, S., Miranda-Angulo, A. L., Wang, H. & Blackshaw, S. Rax-CreERT2 knock-in mice: a tool for selective and conditional
gene deletion in progenitor cells and radial glia of the retina and hypothalamus. _PLoS One_ 9, e90381 (2014). Article PubMed PubMed Central Google Scholar * Kutsia, M., Takeuchi, Y.,
Ranawat, N. & Masai, I. Single-cell transcriptome analysis reveals heterogeneity and a dynamic regenerative response of quiescent radial glia in adult zebrafish brain. _bioRxiv_,
2022.2007.2027.501663 (2022). * Ryu, S. et al. Orthopedia homeodomain protein is essential for diencephalic dopaminergic neuron development. _Curr. Biol._ 17, 873–880 (2007). Article CAS
PubMed Google Scholar * Bornstein, S. R. et al. Stress-inducible-stem cells: a new view on endocrine, metabolic and mental disease? _Mol. Psychiatry_ 24, 2–9 (2019). Article CAS PubMed
Google Scholar * Bornstein, S. R., Berger, I. & Steenblock, C. Are Nestin-positive cells responsive to stress? _Stress_ 23, 662–666 (2020). Article CAS PubMed Google Scholar *
Ninomiya, E. et al. Glucocorticoids promote neural progenitor cell proliferation derived from human induced pluripotent stem cells. _Springerplus_ 3, 527 (2014). Article PubMed PubMed
Central Google Scholar * Heidt, T. et al. Chronic variable stress activates hematopoietic stem cells. _Nat. Med._ 20, 754–758 (2014). Article CAS PubMed PubMed Central Google Scholar
* Leung, C. S. et al. Chronic stress-driven glucocorticoid receptor activation programs key cell phenotypes and functional epigenomic patterns in human fibroblasts. _iScience_ 25, 104960
(2022). Article CAS PubMed PubMed Central Google Scholar * Nürnberg, E., Horschitz, S., Schloss, P. & Meyer-Lindenberg, A. Basal glucocorticoid receptor activation induces
proliferation and inhibits neuronal differentiation of human induced pluripotent stem cell-derived neuronal precursor cells. _J. Steroid Biochem. Mol. Biol._ 182, 119–126 (2018). Article
PubMed Google Scholar * Yu, S. & He, J. Stochastic cell-cycle entry and cell-state-dependent fate outputs of injury-reactivated tectal radial glia in zebrafish. _eLife_ 8, e48660
(2019). Article CAS PubMed PubMed Central Google Scholar * Noorlander, C. W. et al. Antenatal glucocorticoid treatment affects hippocampal development in mice. _PLoS One_ 9, e85671
(2014). Article PubMed PubMed Central Google Scholar * Loi, M., Koricka, S., Lucassen, P. J. & Joels, M. Age- and sex-dependent effects of early life stress on hippocampal
neurogenesis. _Front Endocrinol._ 5, 13 (2014). Article Google Scholar * Best, C., Kurrasch, D. M. & Vijayan, M. M. Maternal cortisol stimulates neurogenesis and affects larval
behaviour in zebrafish. _Sci. Rep._ 7, 40905 (2017). Article CAS PubMed PubMed Central Google Scholar * Shainer, I. et al. Agouti-related protein 2 is a new player in the teleost stress
response system. _Curr. Biol._ 29, 2009–2019.e2007 (2019). Article CAS PubMed PubMed Central Google Scholar * Wasserman-Bartov, T. et al. Tsh induces Agrp1 neuron proliferation in
Oatp1c1-deficient Zebrafish. _J. Neurosci._ 42, 8214–8224 (2022). Article CAS PubMed PubMed Central Google Scholar * Yokobori, E. et al. Stimulatory effect of intracerebroventricular
administration of orexin A on food intake in the zebrafish, Danio rerio. _Peptides_ 32, 1357–1362 (2011). Article CAS PubMed Google Scholar * Smith, K. L. et al. Overexpression of CART
in the PVN increases food intake and weight gain in rats. _Obesity_ 16, 2239–2244 (2008). Article CAS PubMed Google Scholar * Berman, J. R., Skariah, G., Maro, G. S., Mignot, E. &
Mourrain, P. Characterization of two melanin-concentrating hormone genes in zebrafish reveals evolutionary and physiological links with the mammalian MCH system. _J. Comp. Neurol._ 517,
695–710 (2009). Article CAS PubMed PubMed Central Google Scholar * Matsuda, K. et al. Central administration of melanin-concentrating hormone (MCH) suppresses food intake, but not
locomotor activity, in the goldfish, Carassius auratus. _Neurosci. Lett._ 399, 259–263 (2006). Article CAS PubMed Google Scholar * Li, S.-Y. et al. CRH neurons in the lateral
hypothalamic area regulate feeding behavior of mice. _Curr. Biol._ 33, 4827–4843.e7 (2023). Article CAS PubMed Google Scholar * Heinrichs, S. C., Menzaghi, F., Pich, E. M., Hauger, R. L.
& Koob, G. F. Corticotropin-releasing factor in the paraventricular nucleus modulates feeding induced by neuropeptide Y. _Brain Res._ 611, 18–24 (1993). Article CAS PubMed Google
Scholar * Zhang, X. & van den Pol, A. N. Hypothalamic arcuate nucleus tyrosine hydroxylase neurons play orexigenic role in energy homeostasis. _Nat. Neurosci._ 19, 1341–1347 (2016).
Article CAS PubMed PubMed Central Google Scholar * Bolborea, M., Pollatzek, E., Benford, H., Sotelo-Hitschfeld, T. & Dale, N. Hypothalamic tanycytes generate acute hyperphagia
through activation of the arcuate neuronal network. _Proc. Natl. Acad. Sci. USA_ 117, 14473–14481 (2020). Article CAS PubMed PubMed Central Google Scholar * Faught, E. & Vijayan, M.
M. Loss of the glucocorticoid receptor in zebrafish improves muscle glucose availability and increases growth. _Am. J. Physiol. Endocrinol. Metab._ 316, E1093–e1104 (2019). Article CAS
PubMed PubMed Central Google Scholar * Dantzer, B. et al. Density triggers maternal hormones that increase adaptive offspring growth in a wild mammal. _Science_ 340, 1215–1217 (2013).
Article CAS PubMed Google Scholar * Metcalfe, N. B. & Monaghan, P. Compensation for a bad start: grow now, pay later? _Trends Ecol. Evol._ 16, 254–260 (2001). Article PubMed Google
Scholar * Lee, W.-S., Monaghan, P. & Metcalfe, N. B. Experimental demonstration of the growth rate–lifespan trade-off. _Proc. R. Soc. B: Biol. Sci._ 280, 20122370 (2013). Article
Google Scholar * Lemaître, J.-F. et al. Early-late life trade-offs and the evolution of ageing in the wild. _Proc. R. Soc. B: Biol. Sci._ 282, 20150209 (2015). Article Google Scholar *
McEwen, B. S. & Wingfield, J. C. The concept of allostasis in biology and biomedicine. _Horm. Behav._ 43, 2–15 (2003). Article PubMed Google Scholar * Monaghan, P. Organismal stress,
telomeres and life histories. _J. Exp. Biol._ 217, 57–66 (2014). Article PubMed Google Scholar * Jennings, B. J., Ozanne, S. E. & Hales, C. N. Nutrition, oxidative damage, telomere
shortening, and cellular senescence: individual or connected agents of aging? _Mol. Genet. Metab._ 71, 32–42 (2000). Article CAS PubMed Google Scholar * Costantini, D., Marasco, V. &
Møller, A. P. A meta-analysis of glucocorticoids as modulators of oxidative stress in vertebrates. _J. Comp. Physiol. B_ 181, 447–456 (2011). CAS PubMed Google Scholar * Pizzino, G. et
al. Oxidative stress: harms and benefits for human health. _Oxid. Med. Cell Longev._ 2017, 8416763 (2017). Article PubMed PubMed Central Google Scholar * Barnes, R. P., Fouquerel, E.
& Opresko, P. L. The impact of oxidative DNA damage and stress on telomere homeostasis. _Mech. Ageing Dev._ 177, 37–45 (2019). Article CAS PubMed Google Scholar * Burraco, P.,
Díaz-Paniagua, C. & Gomez-Mestre, I. Different effects of accelerated development and enhanced growth on oxidative stress and telomere shortening in amphibian larvae. _Sci. Rep._ 7, 7494
(2017). Article PubMed PubMed Central Google Scholar * Kumari, R. & Jat, P. Mechanisms of cellular senescence: cell cycle arrest and senescence associated secretory phenotype.
_Front. Cell Dev. Biol._ 9, 645593 (2021). Article PubMed PubMed Central Google Scholar * Yang, F. et al. Secretory galectin-3 induced by glucocorticoid stress triggers stemness
exhaustion of hepatic progenitor cells. _J. Biol. Chem._ 295, 16852–16862 (2020). Article CAS PubMed Google Scholar * Bose, R. et al. Glucocorticoids induce long-lasting effects in
neural stem cells resulting in senescence-related alterations. _Cell Death Dis._ 1, e92 (2010). Article CAS PubMed PubMed Central Google Scholar * Anchelin, M. et al. Premature aging in
telomerase-deficient zebrafish. _Dis. Model Mech._ 6, 1101–1112 (2013). CAS PubMed PubMed Central Google Scholar * Bobba-Alves, N. et al. Cellular allostatic load is linked to increased
energy expenditure and accelerated biological aging. _Psychoneuroendocrinology_ 155, 106322 (2023). * Bacila, I., Cunliffe, V. T. & Krone, N. P. Interrenal development and function in
zebrafish. _Mol. Cell. Endocrinol._ 535, 111372 (2021). Article CAS PubMed Google Scholar * Pippal, J. B., Cheung, C. M. I., Yao, Y.-Z., Brennan, F. E. & Fuller, P. J.
Characterization of the zebrafish (Danio rerio) mineralocorticoid receptor. _Mol. Cell. Endocrinol._ 332, 58–66 (2011). Article CAS PubMed Google Scholar * Reul, J. M., de & Kloet,
E. R. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. _Endocrinology_ 117, 2505–2511 (1985). Article CAS PubMed Google Scholar *
Munier, M. et al. Regulation of mineralocorticoid receptor expression during neuronal differentiation of murine embryonic stem cells. _Endocrinology_ 151, 2244–2254 (2010). Article CAS
PubMed Google Scholar * Viho, E. M. G., Buurstede, J. C., Berkhout, J. B., Mahfouz, A. & Meijer, O. C. Cell type specificity of glucocorticoid signaling in the adult mouse hippocampus.
_J. Neuroendocrinol._ 34, e13072 (2022). Article CAS PubMed Google Scholar * Munier, M., Law, F., Meduri, G., Le Menuet, D. & Lombès, M. Mineralocorticoid receptor overexpression
facilitates differentiation and promotes survival of embryonic stem cell-derived neurons. _Endocrinology_ 153, 1330–1340 (2012). Article CAS PubMed Google Scholar * McCann, K. E. et al.
Novel role for mineralocorticoid receptors in control of a neuronal phenotype. _Mol. Psychiatry_ 26, 350–364 (2021). Article CAS PubMed Google Scholar * Schouten, M. et al. Circadian
glucocorticoid oscillations preserve a population of adult hippocampal neural stem cells in the aging brain. _Mol. Psychiatry_ 25, 1382–1405 (2020). Article CAS PubMed Google Scholar *
Zannas, A. S. et al. Epigenetic upregulation of FKBP5 by aging and stress contributes to NF-κB–driven inflammation and cardiovascular risk. _Proc. Natl. Acad. Sci._ 116, 11370–11379 (2019).
Article CAS PubMed PubMed Central Google Scholar * Prekovic, S. et al. Glucocorticoid receptor triggers a reversible drug-tolerant dormancy state with acquired therapeutic
vulnerabilities in lung cancer. _Nat. Commun._ 12, 4360 (2021). Article CAS PubMed PubMed Central Google Scholar * Westerfield M. _The zebrafish book. A guide for the laboratory use of
zebrafish (Danio rerio)_, 4th edn, (University of Oregon Press, 2000). * Choi, M-K. et al. Comprehensive analysis methods for developmental GC exposed zebrafish. Preprint at
https://doi.org/10.17504/protocols.io.kxygx9ooog8j/v3 (2023). * Eachus, H. et al. Disrupted-in-Schizophrenia-1 is essential for normal hypothalamic-pituitary-interrenal (HPI) axis function.
_Hum. Mol. Genet_ 26, 1992–2005 (2017). Article CAS PubMed PubMed Central Google Scholar * Herzog, W. et al. Adenohypophysis formation in the zebrafish and its dependence on sonic
hedgehog. _Dev. Biol._ 254, 36–49 (2003). Article CAS PubMed Google Scholar * Inoue, D. & Wittbrodt, J. One for all–a highly efficient and versatile method for fluorescent
immunostaining in fish embryos. _PLoS One_ 6, e19713 (2011). Article CAS PubMed PubMed Central Google Scholar * Wolf, A. & Ryu, S. Specification of posterior hypothalamic neurons
requires coordinated activities of Fezf2, Otp, Sim1a and Foxb1.2. _Development_ 140, 1762–1773 (2013). Article CAS PubMed Google Scholar * Randlett, O. et al. Whole-brain activity
mapping onto a zebrafish brain atlas. _Nat. Methods_ 12, 1039–1046 (2015). Article CAS PubMed PubMed Central Google Scholar * Liu, N.-A. et al. Pituitary corticotroph ontogeny and
regulation in transgenic Zebrafish. _Mol. Endocrinol._ 17, 959–966 (2003). Article CAS PubMed Google Scholar * Ziv, L. et al. An affective disorder in zebrafish with mutation of the
glucocorticoid receptor. _Mol. Psychiatry_ 18, 681–691 (2013). Article CAS PubMed Google Scholar * Lee, H. B. et al. Novel zebrafish behavioral assay to identify modifiers of the rapid,
nongenomic stress response. _Genes Brain Behav._ 18, e12549 (2019). Article PubMed Google Scholar * Idilli, A. I. et al. Expression of tert Prevents ALT in Zebrafish Brain Tumors. _Front.
Cell Dev. Biol._ 8, 65 (2020). Article PubMed PubMed Central Google Scholar * Eachus, H., Choi, M. K. & Ryu, S. The effects of early life stress on the brain and behaviour: insights
from Zebrafish Models. _Front. Cell Dev. Biol._ 9, 657591 (2021). Article PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS This project is supported by an award
from The Dennis and Mireille Gillings Foundation to S.R. and the German Federal Office for Education Research (grant number 01GQ1404) to S.R. H.E. received support from the Society for
Endocrinology and Wellcome Trust Institutional Strategic Support Fund 3 scheme (ISSF3) to Translational Research Exchange @ Exeter. M.H. received support from the CN Yang Scholars Programme
at Nanyang Technological University. We would like to acknowledge the support of Ms. Kathrin Domdera at the University of Mainz and the Exeter Aquatic Resources Centre staff at the
University of Exeter for expert zebrafish care as well as Dr Corin Liddle at Exeter Bioimaging Centre for microscopy. We are grateful to Dr Elina Jacobs, Dr Kate Ellacott, Dr Steffen
Scholpp, and Professor Gil Levkowitz for feedback on the manuscript. We are grateful to Dr Sara Kuntz for the generation of the _Tg(star:bPAC-2A-tdTomato)__uex300_ transgenic line. AUTHOR
INFORMATION Author notes * Helen Eachus Present address: Institute of Health and Neurodevelopment & Aston Pharmacy School, Aston University, Birmingham, B4 7ET, UK AUTHORS AND
AFFILIATIONS * Living Systems Institute & Department of Clinical and Biomedical Sciences, University of Exeter, Stocker Road, Exeter, EX4 4QD, UK Helen Eachus, Min-Kyeung Choi, Anna
Tochwin & Soojin Ryu * Institute of Human Genetics, University Medical Center, Johannes Gutenberg University Mainz, Langenbeckstraße 1, 55131, Mainz, Germany Johanna Kaspareit * Nanyang
Technological University, 50 Nanyang Avenue, Singapore, 639798, Singapore May Ho Authors * Helen Eachus View author publications You can also search for this author inPubMed Google Scholar *
Min-Kyeung Choi View author publications You can also search for this author inPubMed Google Scholar * Anna Tochwin View author publications You can also search for this author inPubMed
Google Scholar * Johanna Kaspareit View author publications You can also search for this author inPubMed Google Scholar * May Ho View author publications You can also search for this author
inPubMed Google Scholar * Soojin Ryu View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS H.E. designed the study, performed the experiments,
analysed the data, provided funding and wrote the manuscript. M.K.C. designed, performed and analysed the RNA-seq experiments, and performed the GR-ChIP. A.T., J.K. and M.H. performed some
of the experiments. S.R. provided funding, contributed to study design, interpretation of the data and manuscript writing. CORRESPONDING AUTHOR Correspondence to Soojin Ryu. ETHICS
DECLARATIONS COMPETING INTERESTS S.R. holds a patent: US patent number 10,080,355: “A novel inducible model of stress.”. The authors declare no competing interests. PEER REVIEW PEER REVIEW
INFORMATION _Communications Biology_ thanks Caroline Wee and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Ivo Lieberam
and David Favero. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY
INFORMATION SUPPLEMENTARY INFORMATION DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES SUPPLEMENTARY DATA 1 SUPPLEMENTARY DATA 2 SUPPLEMENTARY VIDEO REPORTING SUMMARY RIGHTS AND PERMISSIONS
OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or
other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in
the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Eachus, H., Choi, MK.,
Tochwin, A. _et al._ Elevated glucocorticoid alters the developmental dynamics of hypothalamic neurogenesis in zebrafish. _Commun Biol_ 7, 416 (2024).
https://doi.org/10.1038/s42003-024-06060-5 Download citation * Received: 10 July 2023 * Accepted: 16 March 2024 * Published: 05 April 2024 * DOI: https://doi.org/10.1038/s42003-024-06060-5
SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to
clipboard Provided by the Springer Nature SharedIt content-sharing initiative