Play all audios:
ABSTRACT The vertical structural complexity (VSC) of plant communities reflects the occupancy of spatial niches and is closely related to resource utilization and environmental adaptation.
However, understanding the large-scale spatial pattern of VSC and its underlying mechanisms remains limited. Here, we systematically investigate 2013 plant communities through grid sampling
on the Tibetan Plateau. VSC is quantified as the maximum plant height within a plot (Height-max), coefficient of variation of plant height (Height-var), and Shannon evenness of plant height
(Height-even). Precipitation dominates the spatial variation in VSC in forests and shrublands, supporting the classic physiological tolerance hypothesis. In contrast, for alpine meadows,
steppes, and desert grasslands in extreme environments, non-resource limiting factors (e.g., wide diurnal temperature ranges and strong winds) dominate VSC variation. Generally, with the
shifting of climate from favorable to extreme, the effect of resource availability gradually decreases, but the effect of non-resource limiting factors gradually increases, and that the
physiological tolerance hypothesis only applicable in favorable conditions. With the help of machine learning models, maps of VSC at 1-km resolution are produced for the Tibetan Plateau. Our
findings and maps of VSC provide insights into macroecological studies, especially for adaptation mechanisms and model optimization. SIMILAR CONTENT BEING VIEWED BY OTHERS FUTURE TREE
SURVIVAL IN EUROPEAN FORESTS DEPENDS ON UNDERSTOREY TREE DIVERSITY Article Open access 01 December 2022 INCREASE IN FOREST STRUCTURAL COMPLEXITY ALONG A PRECIPITATION GRADIENT IS MEDIATED BY
PARTIAL HARVESTS IN TEMPERATE PATAGONIAN FORESTS Article Open access 13 June 2024 INTEGRATING FIELD- AND REMOTE SENSING DATA TO PERCEIVE SPECIES HETEROGENEITY ACROSS A CLIMATE GRADIENT
Article Open access 02 January 2024 INTRODUCTION Vertical structural complexity (VSC) in plant heights, which is physical niche partitioning in above-ground space, is thought to be an
important property for a specific plant community1. VSC is closely linked to various ecological processes1,2. For example, in research on the diversity-productivity relationships, the
classical complementary effect3 proposes that the spatial complementarity due to VSC is a key determinant of overyielding in species-rich communities4,5,6, as more complex vertical
structures mean stronger spatial niche partitioning, thereby reducing competition7,8 and leading to the unique occupancy of niche axes such as light2,4. However, few large-scale studies
provide direct evidence that VSC links the diversity-productivity relationship because VSC is rarely quantified. We have conducted a statistical analysis of 136 documents from various
locations worldwide, and only 10 of them have quantified VSC (Table S1). In addition, a recent study showed that VSC has a stronger ability to explain spatial variation in productivity than
species diversity at regional scales9. Hence, it is imperative to investigate the underlying mechanisms of VSC spatial variation and develop high-resolution VSC atlases, which will aid in
understanding the diversity-productivity relationship and accurately predicting spatial variations in productivity. The knowledge of the spatial variation of VSC at large scale is mainly
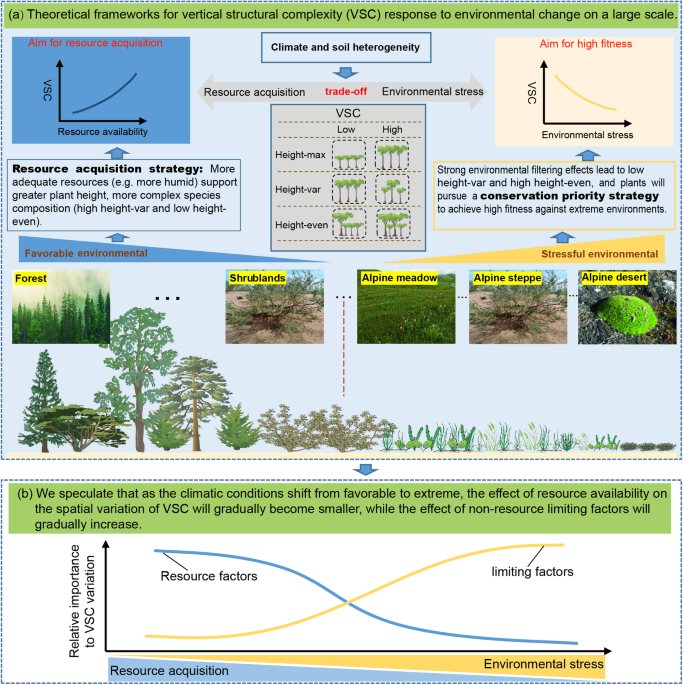
explained by the physiological tolerance hypothesis (PTH)10,11,12. PHT reveals VSC spatial variation mainly from the perspective of regional differences in resource availability9. The PTH
proposes that more adequate resources (e.g., more humid and warmer climates) support greater plant height, more complex species composition, and a wider spectrum of plant functional
strategies (for example, greater crown plasticity and more shade-tolerant species), resulting in greater VSC10,11. Differently, related studies at the local community scale have more often
attributed the complex vertical structure to asymmetric competition between plants. Competition often results in inconsistent individual sizes and promotes complex vertical structures,
because larger individuals gain more resources per unit of biomass and should inhibit the growth of their smaller neighbors13. In addition, the intensity of competition between species will
be greater in areas with superior environments14. The resource acquisition strategies (resource availability and competition for resources) highlighted by PTH and asymmetric competition help
to some extent the understanding of VSC spatial variation10,15. However, a trade-off exists among the plant species in communities to acquire resources for rapid growth in “favorable
conditions” (when resource availability is high and environmental stresses are low) _vs_. its ability to acquire higher fitness to avoid mortality in “extreme conditions” (when resource
availability is low and environmental stresses are high)14,16,17,18. Therefore, we assumed that as climatic conditions shift from favorable to extreme, the impact of resource availability on
spatial variation of VSC will decrease as environmental stress increases, while the effect size of non-resource limiting factors (that is, environmental factors that cause physiological
restrictions on plant growth and reproduction without providing resources) such as extreme low temperatures, high ultraviolet radiation, and strong wind will gradually increase (Fig. 1).
When resource availability is higher (e.g., favorable precipitation and heat conditions), the resource acquisition strategy of plants dominates VSC variation (emphasized by PTH). When
environmental stresses are high, the environmental filtering effects of non-resource limiting factors will be strong, implying a simpler community composition under harsher environmental
conditions19,20,21. In addition, plants that have adapted to harsh environments should pursue a conservation strategy (e.g., morphological changes in plants and facilitations) to avoid
mortality16,22. For example, under conditions of extremely low temperatures and strong winds, plants will maintain a suitable temperature in their immediate environment by growing near other
plants and growing closer to the ground, which is similar to the idea of “warming in a group”23. This dwarf plant community avoids freezing damage caused by low temperatures and
wind-shearing forces caused by strong winds23,24. Although these theoretical underpinnings provide the basis for our extrapolations, but few empirical studies have been able to span from the
favorable to extreme climate gradient. The Tibetan Plateau is an ideal region for investigating the spatial patterns and underlying mechanisms of VSC, because it has a wide range of biomes
from subtropical forests to tundra, a consequence of the variation in environments resulting from the broad altitudinal range in the region (Fig. 2b). Considering this variation, it is also
an important ecological reserve and has always been a hotspot for ecologists. We surveyed 2013 standard plots using a grid-sampling method, covering five major vegetation types in the region
(Fig. 2a). Three parameters were selected to quantify VSC, viz. maximum plant height within a plot (Height-max), the coefficient of variation of plant height (Height-var), and the Shannon
evenness of plant height (Height-even). These three parameters can fully describe the spatial niche occupancy of community individuals in the vertical dimension2. The main objectives of this
study were to: (1) explore the changes in VSC of different vegetation types on the Tibetan Plateau; (2) identify the factors influencing VSC spatial variation; and (3) produce 1-km spatial
resolution maps of VSC, which could provide important parameters for related macroecological studies. Specifically, we tested our assumption that the underlying mechanisms that dominate VSC
variation will gradually shift from resource acquisition strategies to conservation priority strategies as environmental conditions shift from favorable to extreme (Fig. 1), and that the
classic PTH would be more applicable in “favorable conditions”. RESULTS CHANGES IN VSC AMONG DIFFERENT VEGETATION TYPES ON THE TIBETAN PLATEAU The Height-max in the forests, shrublands,
alpine meadows, alpine steppes and alpine desert grasslands were 23.48 (±8.11 SD), 1.23 (±0.51 SD), 0.17 (±0.11 SD), 0.16 (±0.08 SD) and 0.17 (±0.10 SD) m, respectively. The Height-var of
the alpine meadows was larger than that of the alpine steppes, followed by the alpine desert grasslands, but the Height-even of the alpine desert grasslands was greater than that of the
alpine steppes, followed by alpine meadows (Table 1). SPATIAL PATTERNS OF VSC ON THE TIBETAN PLATEAU The Height-max decreases with latitude gradient, and communities growing between 26‒29° N
were 93 times taller than those growing between 38‒41° N (Fig. 3). The Height-max increased and then decreased with the altitude gradient and was the tallest at 1000‒2000 m. Furthermore,
the mean Height-var was smaller between 38‒41° N and 5000‒6000 m than elsewhere. No clear pattern of variation in Height-even along latitude and elevation was observed (Fig. 3). From the
southeast to the northwest of the Tibetan Plateau, the Height-max decreases while the Height-even increases (Fig. 4a, c). The Height-var was lowest in the central regions of the Tibetan
Plateau (Fig. 4b). ENVIRONMENTAL FACTORS INFLUENCING VSC For Height-max and Height-var, aridity index was the most influential factor in forests and shrublands (Table 2) and increased with
precipitation availability (Fig. S1). However, in alpine grassland ecosystems (including alpine meadows, alpine steppes, and alpine desert grasslands), the non-resource limiting factors
(especially diurnal temperature range [Tdiurnal], wind, and oxygen partial pressure [PO2]) played a more important role in the spatial variation of Height-max and Height-var (Table 2). The
stronger the environmental stress, the smaller the Height-max and Height-var (Fig. S1). For Height-even, Aridity index was the dominant factor in shrublands (R2 = 0.669), and wind explained
11.0% of the variation in alpine meadows. However, in the analysis of forests, alpine steppes, and alpine desert grasslands, the explanatory power of a single variable for Height-even
variation was low (less than 10%) (Table 2). Furthermore, the spatial autocorrelation value of the model residuals was very close to zero, indicating that the effect of spatial
autocorrelation on the multiple stepwise regression results was negligible (Fig. S2). Furthermore, the results of the partial correlation analysis showed that the zero-order correlation
coefficient was not significantly different from the partial correlation coefficient, indicating that the grassland community sampling method had a negligible effect (Table S2). POTENTIAL
MECHANISMS FOR THE SPATIAL VARIATION IN VSC Summary based on multiple regression in Table 2 (original samples) and Table S3 (random samples), when climatic conditions gradually change from
favorable to extreme (transition from forest to alpine grassland ecosystem), the effects of resource availability on the variation in VSC were gradually declined from subtropical forests to
alpine desert grasslands, and the effects of non-resource limiting factors have become larger (Fig. 5). HIGH-RESOLUTION MAPS OF VSC ON THE TIBETAN PLATEAU Based on machine learning models,
maps of Height-max, Height-var, and Height-even with 1-km resolution were first produced on the Tibetan Plateau (Fig. 4). The models explained 85%, 55%, and 33% of the spatial variations in
Height-max, Height-var, and Height-even, respectively (Fig. S3). Annual temperature range (Tannual) was the most important predictor of LogHeight-max spatial variation, followed by oil
organic carbon (SOC) and Tdiurnal. Wind was the most important predictor of Height-var spatial variation, followed by UR and minimum temperature of the coldest month (Tcoldest). Wind was the
most important predictor of Height-even spatial variation, followed by mean annual precipitation (MAP) and ultraviolet radiation (UR). To test the model, 25% of the randomly selected plot
data were used. The model predictions of Height-max, Height-var and, Height-even were accurate, because the observed and predicted values were closely and evenly distributed on both sides of
the 1:1 line (Fig. 4). DISCUSSION Based on large-scale field survey data on the Tibetan Plateau, this study explains the determining mechanism of spatial variation in VSC, and produces 1-km
resolution spatial atlas based on machine learning models. We found that Height-max decreases with latitude, which is consistent with the global study by Moles, et al. (2016)25, and further
deepens our knowledge that plants are smaller at high latitudes26. Plant height first increased and then decreased along the altitudinal gradient, which is inconsistent with the
monotonically decreasing trend found by Mao, et al. (2016)27 on the Tibetan Plateau, and is likely due to optimal precipitation and temperature conditions at mid-altitude regions28. We also
found that Height-var was lowest in the regions of maximum altitude (5000‒6000 m) and highest latitude (38‒41°N). These findings confirmed our inference that plant communities possess
shorter and more uniform vertical structures under extreme environmental conditions. We found that aridity index was the dominant factor in the spatial variation of Height-max and Height-var
in forest and shrubland ecosystems, and wetter environments supported higher and more complex community structures (Table 2, Fig. S1). These findings support classic PTH10,11,29. These
results highlight the importance of considering future changes in water availability in forest and shrubland ecosystems, with predictions that the forest and shrubland communities in the
eastern part of the Tibetan Plateau will become taller and more complex under wetter and warmer conditions in the Tibetan Plateau30. As expected, resource availability had weak explanatory
power in alpine ecosystems, such as alpine meadows, alpine steppes, and alpine desert grasslands, where the influence of non-resource limiting factors became stronger (Table 2, Fig. 5). For
the alpine steppes and alpine desert grasslands on the Tibetan Plateau, the most influential factor on Height-max was Tdiurnal (Table 2). In the context of global warming, we predict that
plants in the alpine steppes and alpine desert grasslands will become taller, as the Tdiurnal will become smaller due to the warmer conditions at night than during the day31, thus reducing
the constraints on plant growth. This is consistent with the prediction by Olson, et al.32 that plant height in the Arctic tundra will increase under global warming. Furthermore, Wind
(non-resource limiting factors) was an important predictor of Height-var and Height-even in alpine meadows, alpine steppes, and alpine deserts (Table 2). In the context of climate change,
there is no consensus on how the wind speed of the Tibetan Plateau will change, and both increases33 and decreases34 have been predicted. Therefore, how the Height-var and Height-even of
alpine grasslands will change in the future is uncertain. Considering a wider climatic gradient and different vegetation types, the spatial variation of VSC should be the result of the
trade-off between acquiring resources and achieving greater fitness to avoid mortality (Figs. 5, 6), which is unlike the predictions based on PTH10,11,12,29. The potential mechanisms of
non-resource limiting factors on VSC spatial variation could be explained as follows. First, the environmental filtering effect selects species that can survive in alpine habitats, which
implies a simpler community composition under harsher environmental conditions10,35. Second, these plant communities are tightly packed and grow close to the ground, thereby reducing their
exposure to extreme climates. This is a survival strategy being similar to “warming in a group”23. Third, under extreme environmental conditions, competition between plants is weakened, and
the facilitation will be enhanced for resistance to adverse environmental conditions; compared with competition, facilitation can tolerate the overlap in spatial niches between individuals
and lead to a more uniform community structure18. We produced 1-km resolution maps of the VSC on the Tibetan Plateau for the first time (Fig. 4). These maps can provide important parameters
for macroecological studies from many perspectives, such as the interpretation of underlying mechanisms of the diversity-productivity relationship, more accurate prediction of productivity
variation, and the modification of carbon cycle models15. VSC has extensive connections with ecosystem processes, so its atlas may play important value in other aspects, and its role needs
to be further explored in the future. Specifically, the general connections between VSC and ecosystem processes are as follows: (i) higher vegetation heights will promote primary
productivity, carbon sequestration, and landscape heat uptake and more complex vertical structures will facilitate community function by enhancing spatial complementarity effects3,32; (ii)
higher and more complex vertical structures can promote the coexistence of species at other trophic levels by providing more ecological niches25; and (iii) complex vertical structures mean
communities have diverse physiological traits (such as shade tolerance, crown plasticity, etc.), which will affect the functional diversity of the community and thus the multifunctionality
of the entire ecosystem25 (Fig. 6). Despite the significant advantages and high prediction ability of these maps (Fig. 4), it is a long method that requires improvement. There are few
limitations of the present study. First, several areas were under-sampled in this study, such as the depopulated zone in the northern region of the Tibetan Plateau, which may have resulted
in some uncertainty in the regional estimate. Second, the machine learning model has the advantage of representing heterogeneously distributed observations, because it contains multiple
predictor variables to represent spatial variation in climatic and soil properties36. However, it is the first to estimate these parameters of the VSC at a regional scale and requires
further development in the future. Furthermore, the prediction for Height-even is relatively weak (R2 = 0.33) (Fig. S3). In addition, plant communities in different successional stages have
different resource utilization and acquisition strategies, however, this study failed to consider this during the model construction process due to lack of data9. More importantly, with
technological innovation, we predict that high-resolution remote sensing and terrestrial LiDAR will enable related research to be carried out on a larger scale and provide more useful
information for responding to global climate change (Fig. 6). METHODS STUDY AREA The Tibetan Plateau is the largest (2.5 × 106 km2) and highest plateau in the world37,38. Its unique
topography creates a distinctively variable climatic gradient that provides excellent conditions for validating our hypotheses. The elevation of the Tibetan Plateau ranged from 80 to 8535 m,
mean annual temperature (MAT) ranged from −23 to 24 °C, and mean annual precipitation (MAP) ranged from 18 to 3200 mm. From southeast to northwest, the precipitation and temperature
conditions gradually declined, and the effect of non-resource limiting abiotic factors (e.g., extreme low temperature, diurnal temperature range [Tdiurnal], UR, oxygen partial pressure
[PO2], and wind speed [Wind]) gradually increased (Fig. S4). The Tibetan Plateau mainly includes five vegetation types that vary with climate: subtropical forests, shrublands, alpine
meadows, alpine steppes, and alpine desert grasslands39. In previous studies, ecologists often divided temperature, precipitation, etc. into multiple environmental levels to explore the
variation in biological responses along environmental gradients40. To make the division in the study ecologically meaningful, segmentation was determined at the boundaries between vegetation
types as far as possible40. Exciting, the Tibetan Plateau has a unique advantage in this regard, as the five vegetation types mentioned above correspond closely to the climatic gradients of
the Tibetan Plateau. From subtropical forests to alpine desert grasslands, non-resource limiting factors such as Tdiurnal and Wind gradually strengthen, and the precipitation and
temperature conditions gradually worsen (Fig. 2c). FIELD SAMPLING Field sampling was conducted during the high-growth period from mid-July to late August in 2018, 2019, and 2020. According
to the latitude and longitude, we divided the entire Tibetan Plateau into 1000 grids of equal area (0.5° × 0.5°). For alpine grasslands (i.e., alpine meadows, alpine steppes, and alpine
desert grasslands), we selected the dominant plant communities via visual observation in each grid and randomly set up three 1 m × 1 m plots for the field investigation. There were 1527
plots for grassland communities (Table 1). We determined the tallest plant in each plot by visual inspection, and the height of the tallest plant in each plot was measured with a steel
ruler. For each species within the plots, we recorded their coverage and then randomly selected three plants of each species to measure their height with a steel ruler (all measurements for
less than three individuals); in total, 19,320 plants were measured. For forest and shrubland ecosystems, three 20 m × 20 m plots were randomly established in each grid, for a total of 486
plots (Forests: 456; shrublands: 30; Table 1). For forests, we recorded all vascular plants in these plots, including trees, shrubs, and herbaceous species. Diameter at breast height (DBH)
was measured for trees with a DBH of ˃3 cm. A telescopic stick was used to measure the tree height, and 17,443 trees were measured. The maximum measurement scale of the telescopic stick is
20 m, and the telescopic stick is marked with measurements accurate to centimeters. For those trees with a height greater than 20 m, we complete the measurement based on climbing the trunk.
Only trees with a DBH > 3 cm were measured, as this is a common standard for recording tree layer-related data in forest community surveys41. For shrublands, we recorded their basal
diameter and height, and 339 plants were measured. Across all vegetation types, a total of 2013 field plots were investigated, and the height of 37,102 plants was used for the analysis. The
area within the white curve in Fig. 2a is the depopulated zone, which was not investigated due to challenges accessing these sites. CALCULATIONS OF VSC PARAMETERS Three parameters were used
to quantify VSC for each plot, where Height-max is the maximum plant height within a plot, and Height-var is the coefficient of variation of the plant height, which is calculated using Eq.
1. The Shannon evenness of plant height (Height-even) was calculated using Eq. 21. The smaller the Height-var and the larger the Height-even, the more uniform the height distribution of the
species in the plant community. $${Height}-{{{{\mathrm{var}}}}}=\frac{{{SD}}_{H}}{{{Mean}}_{H}}$$ (1) $${Height}-{even}=\left[-\mathop{\sum }\limits_{k=1}^{{Nh}}{Pk}\times
{{{{\mathrm{ln}}}}}({Pk})\right]/{{{{\mathrm{ln}}}}}({Nh})$$ (2) where _SD__H_ is the standard deviation of plant heights, _Mean__H_ is the mean plant height, _Nh_ number of height classes,
and we used class widths of 100 cm, 10 cm, and 1 cm for forests, shrublands, and grasslands, respectively. For forests and shrublands, _Pk_ refers to the proportion of the basal area for the
kth height class. For grasslands, we divided _Nh_ by the mean height of each species, and _Pk_ refers to the mean of the relative height and relative coverage of the kth height class.
SELECTED EXPLANATORY VARIABLES Based on the research objectives and hypotheses, 13 explanatory variables in total, representing resource- and non-resource limiting variables, were selected.
Resource variables are characterized as precipitation conditions, heat conditions, and variables related to soil fertility, as they are often used to reflect an area’s capacity and resources
for plant growth, reproduction, and survival10,35,42. We chose MAP and an aridity index (AI) to represent precipitation conditions, while MAT represents heat conditions. We measured pH,
soil organic carbon (SOC), and total nitrogen (TN) levels. SOC and TN are important indicators of soil fertility. Soil pH was considered because highly acidic soil affects community species
diversity and regulates soil nutrient supply and uptake by plants42. Growing-season climate variables were not considered, because it is difficult to define a consistent growing season over
such a large spatial scale. Photosynthetically active radiation was also not considered, although competition for light resources within local communities is an important source of VSC. It
has not been considered a resource factor in large-scale studies, because plants only use a very small part of the photosynthetically active radiation reaching the Earth’s surface, generally
considered to be less than 1%20,43. Non-resource limiting variables limit plant growth and reproduction19. As the third pole of the world, alpine ecosystems in the Tibetan Plateau’s central
and northwest regions are usually adapted to stressful environmental conditions. Based on prior knowledge14,21,23,44, we chose the following seven variables: minimum temperature of the
coldest month (Tcoldest), diurnal temperature range (Tdiurnal), annual temperature range (Tannual), atmospheric oxygen partial pressure (PO2), atmospheric carbon dioxide partial pressure
(PCO2), ultraviolet radiation (UR), and Wind. One topsoil sample (0–10 cm) was collected using an auger in each 1 m2 grassland plot. Further, 15‒30 topsoil samples were randomly sampled from
each 400 m2 forest and shrubland plot and then mixed into a composite sample45. The soil samples were air-dried and sieved (<2 mm) to homogenize them. Soil pH was measured using a pH
electrode (Leici), SOC content was measured using the H2SO4-K2Cr2O7 oxidation method, and TN was measured using Kjeldahl nitrogen determination. MAT, MAP, Tcoldest, Tdiurnal, Tannual, and
Wind data were downloaded from the WorldClim database (www.worldclim.org) at a spatial resolution of 30 arc-sec. The mean annual potential evapotranspiration (PET), with a spatial resolution
of 30 arc-sec, was obtained from the Consortium of International Agricultural Research Centers (http://www.cgiar-csi.org/). AI is the ratio of MAP to PET46. Annual average UR was obtained
from the Science Data Bank (https://doi.org/10.11922/sciencedb.332). PO2 and PCO2 were calculated according to the method of Kouwenberg, et al.47. STATISTICS AND REPRODUCIBILITY A machine
learning model (random forest model) was used to upscale the site-level VSC to the entire Tibetan Plateau under contemporary climate scenarios. The predictors included all 13 explanatory
variables considered in this study. We used 75% of the total data as the training data and the remaining 25% as the validation data. The increase in node purity of the splitting variables
was used to estimate the relative importance of the explanatory variables. Based on the spatially gridded data of the predictors (3,617,619 values), the spatial distributions of the VSC for
each grid were mapped at a resolution of 1 × 1 km across the Tibetan Plateau. We interpolated the pH, SOC, TN, PO2, and PCO2 at a spatial resolution of 1-km using “ordinary Kriging
interpolation,” which has the advantage of accurate interpolations at sampling locations48. Glaciers and lakes were removed from the maps. The Height-max was log-transformed to avoid the
influence of data dispersion. The analyses were conducted using MATLAB 2018b (MathWorks, Natick, MA, USA) and the ESRI ArcGIS software (Version 10.2; Redlands, CA, USA). Multiple stepwise
regression (MSR) was used to determine the minimal adequate model, and the “calc.relimp” function in the R package _relaimpo_ was used to estimate the relative importance (R2 and relative
importance metrics[%]) of the two types of variables in the model49. We performed an MSR for the five vegetation types. From subtropical forests to alpine desert grasslands, if the relative
importance of resource variables gradually decreases and non-resource limiting variables increase, our assumption is supported (that is, the underlying mechanisms that dominate VSC variation
will gradually shift from resource acquisition strategies to conservation priority strategies). Considering the differences in sample size may have an impact on the results. We randomly
sampled the four vegetation types using the bootstrapping method and then repeated the above analysis. The shrublands were not considered in this process because it had only 30 plots. The
sample size randomly selected was based on the sample size of the alpine desert grasslands (237 plots), because it has the smallest sample size of the remaining four vegetation types (Table
1). When conducting MSR analysis, we screened explanatory variables based on the following principles. We further screened the 13 explanatory variables based on the Pearson correlation
coefficient, because multicollinearity may affect the determination of the relative importance of factors. Five resource variables (MAT, AI, TN, SOC, and pH) and four non-resource limiting
factors (Tdiurnal, UR, wind, and PO2) were screened. MAP was not considered, because its correlation with MAT (Pearson r = 0.62) was higher than that with AI and MAT (Pearson r = 0.45), and
AI was strongly correlated with MAP (Pearson r = 0.95) (Fig. S1). We only retained Tdiurnal for low temperature-related variables, because it had the lowest correlation with MAT (Pearson r =
−0.46) among all low temperature-related variables, and a strong correlation with Tannual and Tcoldest (Pearson r = −0.71 and −0.60). PCO2 was excluded because it was highly collinear with
PO2 (Pearson’s r = 1). We further employed variance inflation factors (VIF) to verify whether the variables in the optimal model obtained by the MSR are independent19. We also calculated
Moran’s I values for both the observed VSC data and the residuals of the MSR models to examine how the spatial autocorrelation in the VSC was explained by the predictor variables50. MSR
analysis was performed using the stepAIC function of the MASS package (http://www.biostathandbook.com/multipleregression.html). Furthermore, we explored the impact of sampling methods on the
results of grassland communities. Instead of measuring the height of each herb in the grasslands, we randomly selected three plants of each species. Therefore, the number and evenness of
species within the communities may affect the analysis of Height-var and Height-even. To quantify the impact of this sampling, we employed partial correlation analysis. It was judged by
whether the correlations (zero-order correlation coefficient) of Height-var, Height-even, and their dominant factors (based on MSR) changed after controlling for the effect of the
Shannon-Wiener index (partial correlation coefficient). The important values (IV) of the Shannon Wiener index were calculated using the following formula51:
$${{{{{\rm{IV}}}}}}=({{{{{\rm{relative}}}}}}\,{{{{{\rm{height}}}}}}+{{{{{\rm{relative}}}}}}\,{{{{{\rm{coverage}}}}}})/2$$ (3) Graphs were plotted using Origin software (version 8.5;
Northampton, MA, USA). Statistical analysis was performed using R52. The significance level was set at _p_ < 0.05. REPORTING SUMMARY Further information on research design is available in
the Nature Portfolio Reporting Summary linked to this article. DATA AVAILABILITY All data for the figures and tables in the article are provided in the Supplementary Data. REFERENCES *
Danescu, A., Albrecht, A. T. & Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. _Oecologia_ 182, 319–333 (2016). Article
PubMed Google Scholar * Yi, X. X. et al. From canopy complementarity to asymmetric competition: the negative relationship between structural diversity and productivity during succession.
_J. Ecol._ 110, 457–465 (2022). Article Google Scholar * Barry, K. E. et al. The future of complementarity: disentangling causes from consequences. _Trends Eco. Evol._ 34, 167–180 (2019).
Article Google Scholar * Williams, L. J., Paquette, A., Cavender-Bares, J., Messier, C. & Reich, P. B. Spatial complementarity in tree crowns explains overyielding in species mixtures.
_Nat. Eco. Evol._ 1, 0063 (2017). Article Google Scholar * Zhang, Y., Chen, H. Y. H. & Coomes, D. Individual size inequality links forest diversity and above-ground biomass. _J.
Ecol._ 103, 1245–1252 (2015). Article Google Scholar * Lorentzen, S., Roscher, C., Schumacher, J., Schulze, E.-D. & Schmid, B. Species richness and identity affect the use of
aboveground space in experimental grasslands. _Perspect. Plant Ecol._ 10, 73–87 (2008). Article Google Scholar * Chen, Y. et al. Positive effects of neighborhood complementarity on tree
growth in a Neotropical forest. _Ecology_ 97, 776–785 (2016). Article PubMed Google Scholar * Kunz, M. et al. Neighbour species richness and local structural variability modulate
aboveground allocation patterns and crown morphology of individual trees. _Ecol. Lett._ 22, 2130–2140 (2019). Article PubMed Google Scholar * LaRue, E. A. et al. Structural diversity as a
reliable and novel predictor for ecosystem productivity. _Front. Ecol. Environ._ 21, 33–39 (2023). Article Google Scholar * Ehbrecht, M. et al. Global patterns and climatic controls of
forest structural complexity. _Nat. Commun._ 12, 519 (2021). Article CAS PubMed PubMed Central Google Scholar * Harrison, S., Spasojevic, M. J. & Li, D. Climate and plant community
diversity in space and time. _P. Natl Acad. Sci. USA_ 117, 4464–4470 (2020). Article CAS Google Scholar * Currie, D. J. et al. Predictions and tests of climate-based hypotheses of
broad-scale variation in taxonomic richness. _Ecol. Lett._ 7, 1121–1134 (2004). Article Google Scholar * Schwinning, S. & Weiner, J. Mechanisms determining the degree of size asymmetry
in competition among plants. _Oecologia_ 113, 447–455 (1998). Article PubMed Google Scholar * Callaway, R. M. et al. Positive interactions among alpine plants increase with stress.
_Nature_ 417, 844–848 (2002). Article CAS PubMed Google Scholar * Tao, S., Guo, Q., Li, C., Wang, Z. & Fang, J. Global patterns and determinants of forest canopy height. _Ecology_
97, 3265–3270 (2016). Article PubMed Google Scholar * Qin, J. et al. Drivers of tree demographic trade-offs in a temperate forest_. Ecosyst._ 9, 100044 (2022). Article Google Scholar *
Wright, S. J. et al. Functional traits and the growth-mortality trade-off in tropical trees. _Ecology_ 91, 3664–3674 (2010). Article PubMed Google Scholar * He, Z., Webster, S. & He,
S. Y. Growth-defense trade-offs in plants. _Curr. Biol._ 32, R634–R639 (2022). Article CAS PubMed Google Scholar * Wang, Z., Fang, J., Tang, Z. & Lin, X. Patterns, determinants and
models of woody plant diversity in China. _Proc. R. Soc. B_ 278, 2122–2132 (2011). Article PubMed Google Scholar * Wang, Z., Tang, Z. & Fang, J. The species–energy hypothesis as a
mechanism for species richness patterns. _Biodivers. Sci._ 17, 613 (2009). Article Google Scholar * Cheng, C. et al. Plant species richness on the Tibetan Plateau: patterns and
determinants. _Ecography_ 2023, e06265 (2022). Article Google Scholar * Zhang, J. et al. Leaf N:P ratio does not predict productivity trends across natural terrestrial ecosystems.
_Ecology_ 103, e3789 (2022). Article PubMed Google Scholar * Ma, L. et al. Comparative proteomic analysis reveals the role of hydrogen sulfide in the adaptation of the alpine plant
Lamiophlomis rotata to altitude gradient in the Northern Tibetan Plateau. _Planta_ 241, 887–906 (2015). Article CAS PubMed Google Scholar * Klein, T., Randin, C. & Korner, C. Water
availability predicts forest canopy height at the global scale. _Ecol. Lett._ 18, 1311–1320 (2015). Article PubMed Google Scholar * Moles, A. T. et al. Global patterns in plant height.
_J. Ecol._ 97, 923–932 (2009). Article Google Scholar * Niklas, K. J., Midgley, J. J. & Rand, R. H. Size-dependent species richness: trends within plant communities and across
latitude. _Ecol. Lett._ 6, 631–636 (2003). Article Google Scholar * Mao, L., Chen, S., Zhang, J. & Zhou, G. Altitudinal patterns of maximum plant height on the Tibetan Plateau. _J.
Plant Ecol._ 11, 85–91 (2018). Google Scholar * McCain, C. M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. _Glob.
Ecol. Biogeogr._ 16, 1–13 (2006). Article Google Scholar * Ryan, M. G. & Yoder, B. J. Hydraulic limits to tree height and tree growth. _BioScience_ 47, 235–242 (1997). Article Google
Scholar * Hao, Z., Ju, Q., Jiang, W. & Zhu, C. Characteristics and scenarios projection of climate change on the Tibetan Plateau. _Sci. World J._ 2013, 129793 (2013). Article Google
Scholar * Ding, Z., Wang, Y. & Lu, R. An analysis of changes in temperature extremes in the Three River Headwaters region of the Tibetan Plateau during 1961–2016. _Atmos. Res._ 209,
103–114 (2018). Article Google Scholar * Olson, M. E. et al. Plant height and hydraulic vulnerability to drought and cold. _P. Natl Acad. Sci. USA_ 115, 7551–7556 (2018). Article CAS
Google Scholar * Wang, H. et al. Some new changes of the regional climate on the Tibetan Plateau since 2000. _Adv. Earth Sci._ 36, 785–796 (2021). Google Scholar * You, Q. et al. Observed
surface wind speed in the Tibetan Plateau since 1980 and its physical causes. _Int. J. Climatol._ 34, 1873–1882 (2014). Article Google Scholar * Francis, A. P. & Currie, D. J. A
globally consistent richness-climate relationship for angiosperms. _Am. Nat._ 161, 523–536 (2003). Article PubMed Google Scholar * Mishra, U. et al. Spatial heterogeneity and
environmental predictors of permafrost region soil organic carbon stocks. _Sci. Adv._ 7, eaaz5236 (2021). Article CAS PubMed PubMed Central Google Scholar * Yao, T. D. et al. The
imbalance of the Asian water tower. _Nat. Rev. Earth Env._ 3, 618–632 (2022). Article Google Scholar * Li, X. et al. Allometry and distribution of nitrogen in natural plant communities of
the Tibetan Plateau. _Front. Plant Sci._ 13, 845813 (2022). Article PubMed PubMed Central Google Scholar * China Vegetation Map Editorial Committee. China Vegetation Atlas. Beijing:
Science Press (2001). * Guo, Y. et al. The community‐level scaling relationship between leaf nitrogen and phosphorus changes with plant growth, climate and nutrient limitation. _J. Ecol._
108, 1276–1286 (2020). Article CAS Google Scholar * Fang, J. et al. The main contents, methods and technical specifications of plant community inventory. _Biodivers. Sci._ 17, 533–548,
(2009). Article Google Scholar * Chen, S. et al. Plant diversity enhances productivity and soil carbon storage. _P. Natl Acad. Sci. USA_ 115, 4027–4032 (2018). Article CAS Google Scholar
* Öpik, H. & Rolfe, S. The Physiology of Flowering Plants. Cambridge University Press, Cambridge, UK. (2005). * Rahbek, C. et al. Humboldt’s enigma: what causes global patterns of
mountain biodiversity? _Science_ 365, 1108–1113 (2019). Article CAS PubMed Google Scholar * Liu, Y. et al. Regional variation in the temperature sensitivity of soil organic matter
decomposition in China’s forests and grasslands. _Glob. Change Biol._ 23, 3393–3402 (2017). Article Google Scholar * Fei, S. et al. Impacts of climate on the biodiversity-productivity
relationship in natural forests. _Nat. Commun._ 9, 5436 (2018). Article CAS PubMed PubMed Central Google Scholar * Kouwenberg, L. L. R., Kurschner, W. M. & McElwain, J. C. Stomatal
frequency change over altitudinal gradients: prospects for paleoaltimetry. _Rev. Mineral. Geochem._ 66, 215–241 (2007). Article CAS Google Scholar * Kreft, H. & Jetz, W. Global
patterns and determinants of vascular plant diversity. _P. Natl Acad. Sci. USA_ 104, 5925–5930 (2007). Article CAS Google Scholar * Grömping, U. Relative importance for linear regression
inR: the packagerelaimpo. _J. Stat. Softw._ 17, 1–27 (2006). Article Google Scholar * Wang, X., Fang, J., Sanders, N. J., White, P. S. & Tang, Z. Relative importance of climate vs
local factors in shaping the regional patterns of forest plant richness across northeast China. _Ecography_ 32, 133–142 (2009). Article CAS Google Scholar * Wang, H., Pang, C., Zhang, F.
& Zhao, C. Interspecific association of dominant species of wetland vegetation in the middle and lower reaches of Fenhe River, Shanxi province of China. _Chin. J. Ecol._ 31, 2507–2512
(2012). Google Scholar * R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria (2021). Download references
ACKNOWLEDGEMENTS This work was supported by the Second Tibetan Plateau Scientific Expedition and Research Program (STEP, 2019QZKK060602), the National Natural Science Foundation of China
[42141004, 42071303, 31988102], and China Science and Technology Cloud. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * State Key Laboratory of Plant Diversity and Specialty Crops, South China
Botanical Garden, Chinese Academy of Sciences, Guangzhou, 510650, China Changjin Cheng * School of Ecology and Nature Conservation, Beijing Forestry University, Beijing, 100083, China
Changjin Cheng * Key Laboratory of Sustainable Forest Ecosystem Management - Ministry of Education, Northeast Forestry University, Harbin, 150040, China Jiahui Zhang & Nianpeng He * Key
Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, 100101, China Mingxu Li,
Congcong Liu & Li Xu * Northeast Asia Ecosystem Carbon Sink Research Center, Northeast Forestry University, Harbin, 150040, China Nianpeng He Authors * Changjin Cheng View author
publications You can also search for this author inPubMed Google Scholar * Jiahui Zhang View author publications You can also search for this author inPubMed Google Scholar * Mingxu Li View
author publications You can also search for this author inPubMed Google Scholar * Congcong Liu View author publications You can also search for this author inPubMed Google Scholar * Li Xu
View author publications You can also search for this author inPubMed Google Scholar * Nianpeng He View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS Nianpeng He designed the research. Changjin Cheng and Jiahui Zhang conducted the research (collected the datasets and analyzed the data). Changjin Cheng wrote the manuscript.
Mingxu Li, Li Xu, and Congcong Liu commented on the details of the manuscript drafts. CORRESPONDING AUTHORS Correspondence to Jiahui Zhang or Nianpeng He. ETHICS DECLARATIONS COMPETING
INTERESTS The authors declare no competing interests. PEER REVIEW PEER REVIEW INFORMATION _Communications Biology_ thanks the anonymous reviewers for their contribution to the peer review of
this work. Primary Handling Editors: Shouli Li and David Favero. A peer review file is available. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature remains neutral with regard to
jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION PEER REVIEW FILE SUPPLEMENTARY INFORMATION DESCRIPTION OF ADDITIONAL SUPPLEMENTARY FILES
SUPPLEMENTARY DATA REPORTING SUMMARY RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing,
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons
licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a
credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted
use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT
THIS ARTICLE CITE THIS ARTICLE Cheng, C., Zhang, J., Li, M. _et al._ Vertical structural complexity of plant communities represents the combined effects of resource acquisition and
environmental stress on the Tibetan Plateau. _Commun Biol_ 7, 395 (2024). https://doi.org/10.1038/s42003-024-06076-x Download citation * Received: 16 March 2023 * Accepted: 19 March 2024 *
Published: 01 April 2024 * DOI: https://doi.org/10.1038/s42003-024-06076-x SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link
Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative