Play all audios:
By maintaining stress factors under optimal low-dose levels, plant performance and productivity can be increased and postharvest produce quality can be improved. Hence, for a sustainable
food production, plant stress should not be feared but systematically monitored and embraced. Numerous chemicals as well as stressors have both favorable and unfavorable consequences for
plant performance and health in a dose-dependent manner, and here we discuss implications of these phenomena for the practice of sustainable agriculture. CROP PRODUCTION IN CHANGING
ENVIRONMENTS Sustainable food production faces an unprecedented crisis facilitated by multifaceted challenges such as toxicities induced in plants by air and soil pollutants at a global
scale, enhanced activities of plant pests and diseases driven by climate change, and development of resistance of pests to pesticides, among others1,2,3,4. Such challenges suppress plant
growth, productivity, and yields, while also downgrading the quality of plant products used for food1,2,3,4. Meanwhile, the population demands for more qualitative foods are increasing,
creating an urgent task to manage plant stress, enhance crop performance and yield, and adapt plants and agroecosystems to global changes5,6. Hence, efficient management of plant stress is
crucial for sustainable food production and the United Nations Sustainable Development Goals achievement in the post-2020 era. However, much is argued and offered as promising solutions by
scientific research over the years, but what can be practically achieved? Potentially applicable and effective solutions at a global scale -including less rich or developed countries- should
be cheaper, less technology-demanding, and easily accessible. For instance, breeding and genetic engineering are profoundly important and offer a unique opportunity for improvements, but
they are time and technology-demanding, and the net benefit such technologies could offer is restricted by the limits of biological plasticity. That is, plants and other organisms have
specific quantitative limits within which their photosynthetic, growth, and yield performance can be increased, and these limits cannot be exceeded. Therefore, instead of applying specific
technologies for a limited benefit, a more universal approach would be to efficiently manage universal plant stress, i.e. the total stress induced in plants by the concurrent exposure to a
broad range of key environmental challenges together in a specific environment. The currently prevailed approach has been to target minimizing factors causing abiotic stress in plants (Table
1) and/or eliminate plant stress. Interventions are also applied to enhance crop productivity under abiotic stress and promote agricultural sustainability, including chemical (e.g.
exogenous chemical application), genetic (e.g. breeding, gene editing), and microbial (e.g. beneficial microorganisms inoculation) approaches7,8. However, the approach of eliminating plant
stress was based on the long-held scientific understanding that stress is harmful to plants and the associated perception stemming from the belief that stress is always bad. This was
primarily due to the longstanding assumption that reactive oxygen species (ROS), which modulate plant stress, are harmful, and that plant traits critical to plant fitness and productivity,
such as photosynthesis and chlorophylls, are suppressed by oxidative stress. Therefore, plant stress should be eliminated and interventions should be applied to increase plant performance
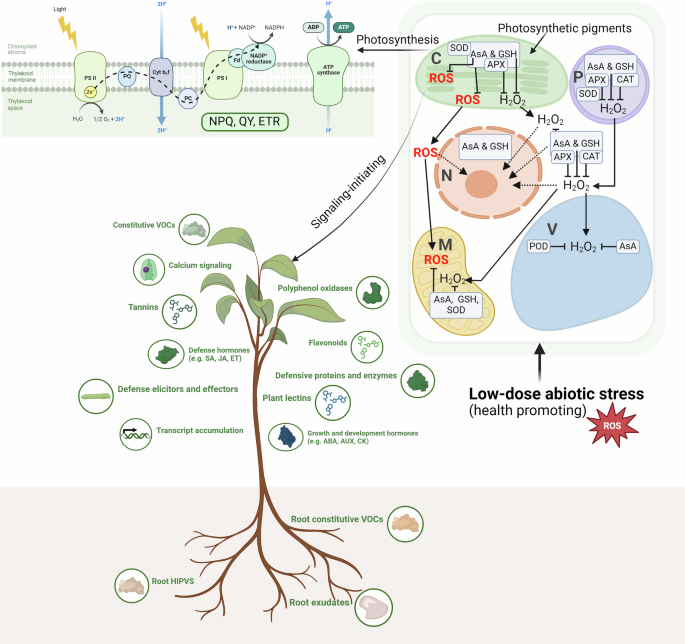
and productivity. WHAT HAS CHANGED IN THE SCIENTIFIC UNDERSTANDING OF PLANT STRESS However, in the last few years, it became clear that ROS (e.g. H2O2) not only are not harmful up to
species-specific thresholds, but they could also be beneficial, acting as secondary messengers to mediate plant immune responses and improve performance and health (Fig. 1)9,10. Hence, ROS
are essential for healthy functioning, and mild increases in ROS induced by stress offer benefits in plants with ROS levels within the optimal range11. This is also true for reactive
nitrogen (e.g. NO) and sulfur (e.g. H2S) species, while there is a known molecular crosstalk among reactive oxygen, nitrogen, and sulfur species11,12. Further, it has recently emerged that
major traits critical to fitness and productivity, such as photosynthesis and chlorophylls, are typically enhanced by doses of stress smaller than the toxicological threshold for damages
(subtoxic doses)10,13,14. The latter low-dose stimulations are linked with the mild ROS increment and associated modulation of the electron transport rate and non-photochemical quenching
(NPQ), which protects the photosynthetic system from damage by excess light13,15,16. Ecophysiological evidence suggests that these mechanisms appear universally across plants and agents
inducing oxidative stress, and act as signals to regulate metabolism and transcription factors and enhance enzymatic and non-enzymatic antioxidants as well as defense hormones (Fig.
1)10,13,14,15,17,18,19. The low-dose stimulation is explained by hormesis, a generalized phenomenon that appears in a vast array of organisms as a biphasic response with restricted low-dose
stimulation of commonly <60% and high-dose inhibition, describing the limits of biological plasticity14,20. Such stimulation by low-dose also preconditions (primes) plants and protects
against more severe stress that may occur subsequently, whether from the same or different biotic or abiotic stressors17,20,21. Therefore, it is now clear that some stress is essential for
improved performance of plants, pointing to the issue of how stress can be better managed to facilitate sustainable food production. However, a question arising is whether the low-dose
stimulation at physiological level may increase fitness in agriculture, promoting sustainable farming that provides resources required to feed the current population while ensuring
sustaining future generations at least as efficiently. In agriculture, besides the enhanced plant physiological performance, an indicator of fitness would be the yields and yield quality,
and recent studies provide support for such desired outcomes. For instance, the herbicide glyphosate increased common bean (_Phaseolus vulgaris_ L.) productivity in both winter season (7.2 g
a.e. ha–1) and wet season (36 and 54 g a.e. ha–1)22. Glyphosate (1.8 g a.e. ha–1) also increased shikimic acid and quinic acid, plant growth, yield, and technological quality of sugarcane
(_Saccharum_ spp.), with enhanced Brix% juice, recoverable sugars, pol% cane, tons of pol ha–1, and tons of culms ha–1 23. Moreover, nickel (Ni) increased photosynthetic pigments and
photosynthesis and ureides and the number of seeds per pot and yield at 0.5 mg Kg−1 in cowpea (_Vigna unguiculata_ L. Walp), whereas it caused adverse effects at 2–3 mg Kg−1 24. Cadmium (Cd)
applied at 0. 5 mg Kg−1 also increased medicinal components (flavonoids) and biomass by ~60% and 20% a medicinal plant (_Polygonatum cyrtonema_ Hua) whose tuber is used in traditional
Chinese medicine19. Similarly, hexavalent chromium (Cr(VI)) application at a low concentration of 0.5 mg L−1 K2Cr2O7 induced hormesis, evident through increased biomass and larger leaves in
alfalfa (_Medicago sativa_ L.) plants, effects that were opposite from those at elevated application (5–10 mg L−1)11. In addition to chemicals, intermittent UV-B exposure (ambient ±7.2 kJ
m−2 d−1) improved the growth of the medicinal plant _Eclipta alba_ L. (Hassak) and the yield of wedelolactone, a promising compound with remarkable pharmacological values25. These selective
examples, including replicated field evaluations22,23, indicate the promise of interventions based on low-dose stimulation to increase productivity, yields, and food quality, aiding to
achieve food security and combat hunger and malnutrition in recognition of the lag behind meeting Sustainable Development Goal (SDG) 2 for ‘Zero Hunger’ 26. However, revealing whether
low-dose stress may increase fitness in agriculture requires integrated cost-benefit analyses in which environmental and economic benefits and impacts, will also be taken into account along
with food safety and other risks (see next section for potential low-dose stimulation of crop pests). IMPLICATIONS FOR PRACTICE It emerges from the preceding discussion that, instead of
applying other technologically demanding interventions to enhance plant performance to a rather limited extent, this could be achieved by producing or allowing an optimum level of stress in
plants, especially since the maximum enhancement is restricted by the limits of biological plasticity and would not be exceeded regardless of the intervention. However, the application of
the concept in the practice is challenging, and there are various aspects that need to be considered in a roadmap toward the implementation of low-dose stress to facilitate sustainable food
production (Fig. 2). It is also important to note that this exposition refers specifically to abiotic stressors as stimulatory agents but does not recommend the use of biotic stressors. That
is because the application of biotic stressors is more challenging, due to population and community ecology phenomena (e.g. network complexity, Allee effect), and might unintentionally
result in pest outbreak or epidemic27. First, substantial evidence emerged in the last few years showing that various chemicals in the environment induce aforementioned positive effects on
many kinds of plants. These include insecticides, fungicides, herbicides, metallic elements, micro/nanoplastics and other particles, human and veterinary pharmaceuticals, and other
chemicals10,13,14,15,18. The dominant stress-inducing chemicals under local conditions should be identified. However, environmental standards and (phyto)remediation programs should consider
targeting not the complete removal of these stressors, but their decrease to concentrations as needed to induce positive effects. For example, a recent meta-analysis of about 8500 dose
responses from the published literature revealed that cerium (Ce), a rare earth element widely applied for its positive effects on plants, commonly increases chlorophylls, stomatal
conductance (uptake of air), photosynthesis, and biomass at concentrations from as low as below 0.1 mg L−1 to 25 mg L−1 but suppresses these traits at concentrations over 50 mg L−1 across
species18. Importantly, current technological advancements make difficult and considerably expensive the removal of the smallest fraction of such chemicals from the environment, perplexing
or making it often practically impossible for the removal of the lowest 10% of the chemicals. Instead, major economic resources and efforts would be saved by allowing the maximum permissible
concentrations in the environment (inducing positive effects on plants), assuming these are below the safety limits for environmental and human safety standards. These new advances in the
scientific understanding of chemical-induced dual (hormetic) effects also indicate that the effects of chemicals widely used in the agricultural practice, as is the case of the long history
of rare earth element application in China, can turn from positive to negative depending on the concentration. Therefore, care should be exercised that major chemicals applied in the field
are maintained at desired levels, avoiding their excessive accumulation and persistence in the environment. To this end, analysis of environmental media every several years (or decadal) to
ensure that the concentrations of few major applied chemicals do not exceed specific thresholds would facilitate maximization of the desired effects on plants. This could be incorporated
into local farming practices, at a relatively low cost, but would require the existence of specialists providing such analytical services. However, in the realm of biodiversity conservation
and SDGs, various types of (hyper)accumulator plants could also be incorporated into the cultivations, for instance as part of a rotation system, in mixed-species cultivation, in specific
sub-plots in a cultivation, and/or surrounding a land plot. This would promote diversification, which facilitates SDG 15 ‘life on land’, while also reducing the concentrations of chemicals
with hormetic effects, such as heavy metals. Routine incorporation of (hyper)accumulators in cropping systems would also provide other benefits, such as potentially reducing crop herbivory
(due to accumulation of chemicals toxic to herbivores), and lowering the need of chemical analyses of environmental media by acting proactively. Important to note is also that abiotic
stress-inducing agents do not need to be environmental contaminants, but in “clean” environments naturally-occurring chemicals and gaseous molecules (e.g. hydrogen peroxide, hydrogen
sulfide, nitric oxide, etc) and/or physical agents with no residue in the environment (e.g. radiation, cold, etc) could be applied under controlled conditions to induce stimulatory adaptive
responses before seedling transplantation into the field. This approach is also highly relevant and applicable to facility agriculture, such as in greenhouses, where stress-free conditions
are commonly targeted. The perspective provided herein indicates that manipulations can easily be applied to create optimum stress conditions. For example, by introducing hormesis-inducing
chemicals and maintaining them at optimum stimulatory concentrations, especially in hydroponic cultures where the regulation is relatively easier and straightforward. Moreover, this concept
shows promise for practical application to induce the production of nutraceuticals and other plant-producing chemicals that are used to improve human health, as low doses of stress boost the
production of such chemicals28. Where plants are cultivated for such use only, plant stress can be maintained up to a level before the nutraceutical levels start decreasing. Besides plants,
such techniques (e.g. low-dose radiation) can be applied to organisms that are beneficial for different purposes in the agricultural practice, such as to promote the growth of silkworms29
or the performance of pollinators30 released to facilitate pollination as for example in the controlled cultivation of some vegetables. Second, the vast majority of scientific research
involves one or at best two abiotic stressors concurrently administered to plants, while in reality there are numerous stressors acting together. It is well-known that the responses of
plants to mixtures of stressors can differ from those of individual stressors, but the low-dose positive effects were widely induced by mixtures, which requires that at least one component
of the mixture inducing stimulatory effects. It is practically infeasible to examine all possible stressors in a given environment, such as a specific crop cultivation area. However,
identifying the top 1–3 abiotic stressors based on their levels and potency and regulating their concentrations at levels expected to induce hormesis may provide a fast-forward solution when
other concurrent stressors are not manipulated. This would require support by local authorities, which would perform the required environmental monitoring and analyses and develop and adopt
relevant policies for the control of the levels of such stressors. Hence, a close cooperation between government actors, policy makers, scientists, industrial stakeholders and end-users
(farmers) is needed. Third, it needs to be understood that, through epigenetic mechanisms, low-dose stimulation of parents can transfer precious information to offspring, which enhances
offspring functioning early or later in their life without the offspring be directly exposed to stress yet17. This conditions offspring to perform better when facing stress later on.
Therefore, the tolerance of plants to stress under local conditions may change over time and generations31, affecting the concentrations needed to induce and maintain stimulatory effects.
This property should be factored in the long-term assessments. Moreover, if low-dose stimulation leads to selection of more resistant individuals within a population as indicated for
weeds32,33, evolutionary agroecology suggests that genotypes with intermediate (but not highest) fitness can produce the highest-yielding populations34. Hence, a target for long-term
assessments would also be to identify the optimum fitness in low dose-stimulated crop populations up to a point after which the yields start to decrease due to exceeding the degree of
fitness that permits highest yields. This natural selection process mediated by low-dose stimulation interventions could further determine the fitness in agriculture and therefore
sustainable crop production. Fourth, there are various ways for detecting plant stress. Among them, chlorophyll fluorescence imaging analysis is a method with great and rapid sensitivity,
detecting stress within 15–30 min15. Importantly, it is non-invasive and relatively low-cost, permitting a wider utilization15. A novel efficient index of early stress detection is the
proportion of open reaction centers of photosystem II (PSII) (q_p_)15, which can be used for systematic monitoring of plant stress status. A question of practical relevance that arises is
how much dose is considered low dose and how a low level of stress can be managed. Accepting that the target is to stimulate cultivated plants, low doses are those that enhance fitness
critical traits (e.g. photosynthesis, chlorophylls, fluorescence quantum yield, q_p_) and endpoints (e.g. growth, survival, productivity, reproduction). A dose to be low (subtoxic) should
not cause biologically negative effects on the cultivated plants, and this can be controlled by monitoring plant health indicators such as quantum yield and q_p_ in order to maintain the
stress at desired levels. A low dose would cause stimulation initially, but the enhancement may not persist, requiring re-application of the stressor. Therefore, weekly observations of
indicators (e.g. quantum yield, q_p_) would reveal whether further stressor addition is needed to extend the stimulation as desired. Considering that plants can acclimate to low-dose stress
and develop tolerance, a higher dose may be needed over time to sustain stimulation. Despite the dose increases, it is still considered a low dose when it causes no biologically negative
effects or even positively affects the cultivated plants. However, if plant stress monitoring indicates tendency for negative effects, the stressor could be minimized or eliminated and allow
plant recovery for 1–2 weeks. Considering also the genotypic degrees of susceptibilities, different cultivars or varieties of the same plant species are expected to have different dose
requirements. Moreover, care should be exercised when selecting doses to apply doses that do not enhance plant pest populations with the potential for outbreak. Therefore, regular
observations of pests are warranted alongside plant stress status monitoring. Fifth, the lowering of concentrations of contaminants in the environment may introduce risks associated with the
low-dose stimulation. For example, recently the European Union proposed to reduce chemical pesticides use and risk by 50% by 2030
(https://food.ec.europa.eu/plants/pesticides/sustainable-use-pesticides_en). However, such a reduction may result in the enhancement of pests, diseases, and weeds of crop cultivations, if
the lower concentrations induce unidentified low-dose stimulation in such biotic threats to crops, as well as the development and promotion of pesticide resistance32,35,36. For example,
herbicide-resistant weeds show enhanced propagation and spread at doses recommended for application to control weeds, and, even in weed populations that are on average sensitive to
herbicides, a subpopulation of ‘stronger’ plants can be favored by hormetic responses32. Moreover, the herbicide glyphosate increased the number of seeds (+114% and +41%) and capitulum
(+37%, +48%) per plant in sensitive (at 0.7 and 2.8 g a.e. ha–1) and resistant (at 5.6 and 5.6 g a.e. ha–1) biotypes of the crop weed _Conyza sumatrensis_31. This indicates increased
reproduction of the weed, and suggests potential risks for fitness in agriculture. Similar findings for hormetic stimulation by such chemicals were reported for other crop weeds, commonly
with resistant biotypes undergoing stimulation by higher doses compared to susceptible biotypes37. Therefore, the low-dose stimulation should be incorporated in all assessments of effects of
and risks introduced by co-occurring organisms that can become harmful to cultivated plants. Sixth, low-dose stimulation by irradiation and other physical and chemical stressors can be
successfully applied in postharvest produce, offering benefits in the secondary metabolites linked with improved human health, facilitating the management of pathogen that downgrade produce
quality, and improving the quality of food products and extending their lifespan38. The biological mechanisms explaining these are in line with those explained for plants, and involve ROS
modulation38. Hence, low-dose stress can decrease postharvest losses, a major undermining factor of food production, and offer remarkable improvement to postharvest produce. All in all, it
is becoming increasingly evident that low-dose stimulation is of vital importance in food production, and its acknowledgement, understanding, inclusion in relevant research, and study can
facilitate sustainable food production. The dual effects of stress present a double-edged sword with various implications for practice, which are summarized herein. REFERENCES *
Agathokleous, E. et al. Adapting crop production to climate change and air pollution at different scales. _Nat. Food_ 4, 854–865 (2023). Article Google Scholar * Ma, C. S. et al. Climate
warming promotes pesticide resistance through expanding overwintering range of a global pest. _Nat. Commun._ 12, 5351 (2021). Article CAS Google Scholar * Singh, B. K. et al. Climate
change impacts on plant pathogens, food security and paths forward. _Nat. Rev. Microbiol._ 21, 640–656 (2023). Article CAS Google Scholar * Deutsch, C. A. et al. Increase in crop losses
to insect pests in a warming climate. _Science (80-.)_ 361, 916–919 (2018). Article CAS Google Scholar * Agathokleous, E., Feng, Z., Frei, M., Jiao, S. & Burkey, K. O. Response and
adaptation of agricultural ecosystems to global changes. _Agric. Ecosyst. Environ._ 362, 108844 (2024). Article Google Scholar * Yoon, D. K. et al. Transgenic rice overproducing Rubisco
exhibits increased yields with improved nitrogen-use efficiency in an experimental paddy field. _Nat. Food_ 1, 134–139 (2020). Article CAS Google Scholar * Zhang, H., Zhu, J., Gong, Z.
& Zhu, J. K. Abiotic stress responses in plants. _Nat. Rev. Genet._ 23, 104–119 (2022). Article Google Scholar * Compant, S. et al. Harnessing the plant microbiome for sustainable crop
production. _Nat. Rev. Microbiol_. https://doi.org/10.1038/s41579-024-01079-1 (2024). * Wu, B., Qi, F. & Liang, Y. Fuels for ROS signaling in plant immunity. _Trends Plant Sci._ 28,
1124–1131 (2023). Article CAS Google Scholar * Jalal, A. et al. Hormesis in plants: physiological and biochemical responses. _Ecotoxicol. Environ. Saf._ 207, 111225 (2021). Article CAS
Google Scholar * Christou, A. et al. Hexavalent chromium leads to differential hormetic or damaging effects in alfalfa (_Medicago sativa_ L.) plants in a concentration-dependent manner by
regulating nitro-oxidative and proline metabolism. _Environ. Pollut._ 267, 115379 (2020). Article CAS Google Scholar * Antoniou, C., Savvides, A., Christou, A. & Fotopoulos, V.
Unravelling chemical priming machinery in plants: the role of reactive oxygen–nitrogen–sulfur species in abiotic stress tolerance enhancement. _Curr. Opin. Plant Biol._ 33, 101–107 (2016).
Article CAS Google Scholar * Moustakas, M., Moustaka, J. & Sperdouli, I. Hormesis in photosystem II: a mechanistic understanding. _Curr. Opin. Toxicol._ 29, 57–64 (2022). Article CAS
Google Scholar * Erofeeva, E. A. Environmental hormesis of non-specific and specific adaptive mechanisms in plants. _Sci. Total Environ._ 804, 150059 (2022). Article CAS Google Scholar
* Moustaka, J. & Moustakas, M. Early-stage detection of biotic and abiotic stress on plants by chlorophyll fluorescence imaging analysis. _Biosensors_ 13, 796 (2023). Article CAS
Google Scholar * Wang, Y. et al. Glyphosate hormesis stimulates tomato (_Solanum lycopersicum_ L.) plant growth and enhances tolerance against environmental abiotic stress by triggering
nonphotochemical quenching. _Pest Manag. Sci._ 80, 3628–3639 (2024). Article CAS Google Scholar * Carvalho, M. E. A., Castro, P. R. C. & Azevedo, R. A. Hormesis in plants under Cd
exposure: from toxic to beneficial element? _J. Hazard. Mater._ 384, 121434 (2020). Article CAS Google Scholar * Agathokleous, E. et al. Mechanisms of cerium-induced stress in plants: a
meta-analysis. _Sci. Total Environ._ 852, 158352 (2022). Article CAS Google Scholar * Yang, L. et al. Unlocking hormesis and toxic effects induced by cadmium in _Polygonatum cyrtonema_
Hua based on morphology, physiology and metabolomics. _J. Hazard. Mater._ 465, 133447 (2024). Article CAS Google Scholar * Erofeeva, E. A. Environmental hormesis: from cell to ecosystem.
_Curr. Opin. Environ. Sci. Heal._ 29, 100378 (2022). Article Google Scholar * Christou, A., Agathokleous, E. & Fotopoulos, V. Safeguarding food security: hormesis-based plant priming
to the rescue. _Curr. Opin. Environ. Sci. Heal._ 28, 100374 (2022). Article Google Scholar * Pacheco de Almeida Prado Bortolheiro, F., Brunelli-Nascentes, M. C., Luiz Santos, H. & de
Almeida Silva, M. Increased hormetic dose of glyphosate causes oxidative stress and reduces yield in common bean. _Plant Stress_ 10, 100231 (2023). Article CAS Google Scholar *
Pincelli-Souza, R. P., Bortolheiro, F. P., Carbonari, C. A., Velini, E. D. & de Silva, M. A. Hormetic effect of glyphosate persists during the entire growth period and increases
sugarcane yield. _Pest Manag. Sci._ 76, 2388–2394 (2020). Article CAS Google Scholar * Mendes, N. A. C. et al. Physiological and biochemical role of nickel in nodulation and biological
nitrogen fixation in _Vigna unguiculata_ L. Walp. _Plant Physiol. Biochem._ 201, 107869 (2023). Article CAS Google Scholar * Rai, K. & Agrawal, S. B. An assessment of dose-dependent
UV-B sensitivity in _Eclipta alba_: biochemical traits, antioxidative properties, and wedelolactone yield. _Environ. Sci. Pollut. Researc_ 28, 45434–45449 (2021). Article CAS Google
Scholar * Sustainable agriculture. _Nat. Sustain_. 1, 531 https://www.nature.com/articles/s41893-018-0163-4, https://doi.org/10.1038/s41893-018-0163-4 (2018). * Guedes, R. N. C., Benelli,
G. & Agathokleous, E. Arthropod outbreaks, stressors and sublethal stress. _Curr. Opin. Environ. Sci. Heal._ 28, 100371 (2022). Article Google Scholar * Caicedo-Lopez, L. H.,
Guevara-Gonzalez, R. G., Ramirez-Jimenez, A. K., Feregrino-Perez, A. A. & Contreras-Medina, L. M. Eustress application trough-controlled elicitation strategies as an effective
agrobiotechnology tool for capsaicinoids increase: a review. _Phytochem. Rev._ 21, 1941–1968 (2022). Article CAS Google Scholar * Shibamoto, Y., Kamei, Y., Kamei, K., Tsuchiya, T. &
Aoyama, N. Continuous low-dose-rate irradiation promotes growth of silkworms. _Dose Res._ 15, 155932581773525 (2017). Article Google Scholar * Ramanaidu, K. & Cutler, G. C. Different
toxic and hormetic responses of _Bombus impatiens_ to _Beauveria bassiana_, _Bacillus subtilis_ and spirotetramat. _Pest Manag. Sci._ 69, 949–954 (2013). Article CAS Google Scholar *
Wang, Y. et al. Glyphosate hormesis effects on the vegetative and reproductive development of glyphosate-susceptible and -resistant _Conyza sumatrensis_ biotypes. _Pest Manag. Sci._ 80,
3628–3639 (2024). Article CAS Google Scholar * Belz, R. G., Carbonari, C. A. & Duke, S. O. The potential influence of hormesis on evolution of resistance to herbicides. _Curr. Opin.
Environ. Sci. Heal._ 27, 100360 (2022). Article Google Scholar * Belz, R. G. & Sinkkonen, A. Low glyphosate doses change reproduction and produce tolerant offspring in dense
populations of _Hordeum vulgare_. _Pest Manag. Sci._ 77, 4770–4784 (2021). Article CAS Google Scholar * Weiner, J., Du, Y. L., Zhang, C., Qin, X. L. & Li, F. M. Evolutionary
agroecology: individual fitness and population yield in wheat (_Triticum aestivum_). _Ecology_ 98, 2261–2266 (2017). Article Google Scholar * Guedes, R. N. C., Smagghe, G., Stark, J. D.
& Desneux, N. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. _Annu. Rev. Entomol._ 61, 43–62 (2016). Article CAS Google Scholar *
Guedes, R. N. C., Rix, R. R. & Cutler, G. C. Pesticide-induced hormesis in arthropods: towards biological systems. _Curr. Opin. Toxicol._ 29, 43–50 (2022). Article CAS Google Scholar
* Anunciato, V. M. et al. Effect of low glyphosate doses on flowering and seed germination of glyphosate-resistant and -susceptible _Digitaria insularis_. _Pest Manag. Sci_. 78, 1227–1239
(2022). * Duarte-Sierra, A., Tiznado-Hernández, M.-E. & Jha, D. K. Postharvest hormesis in produce. _Curr. Opin. Environ. Sci. Heal._ 29, 100376 (2022). Article Google Scholar * Shang,
B. et al. Drought mitigates the adverse effects of O3 on plant photosynthesis rather than growth: a global meta‐analysis considering plant functional types. _Plant. Cell Environ._ 47,
1269–1284 (2024). Article CAS Google Scholar * An, Q., Wen, C. & Yan, C. Meta-analysis reveals the combined effects of microplastics and heavy metal on plants. _J. Hazard. Mater._
476, 135028 (2024). Article CAS Google Scholar * Wang, C. et al. Toxic effects of microplastics and nanoplastics on plants: a global meta-analysis. _Environ. Pollut._ 337, 122593 (2023).
Article CAS Google Scholar * Sun, Y., Wang, C., Chen, H. Y. H. & Ruan, H. Response of plants to water stress: a meta-analysis. _Front. Plant Sci._ 11, 978 (2020). Article Google
Scholar * Feng, Z., Wang, S., Szantoi, Z., Chen, S. & Wang, X. Protection of plants from ambient ozone by applications of ethylenediurea (EDU): a meta-analytic review. _Environ.
Pollut._ 158, 3236–3242 (2010). Article CAS Google Scholar * Zhao, Y. et al. A meta-analysis of elevated O3 effects on herbaceous plants antioxidant oxidase activity. _PLoS One_ 19,
e0305688 (2024). Article Google Scholar Download references ACKNOWLEDGEMENTS The work of E.A. was supported by the National Natural Science Foundation of China (No. 31950410547). AUTHOR
INFORMATION AUTHORS AND AFFILIATIONS * Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters (CIC-FEMD), Nanjing University of Information Science &
Technology, Nanjing, 210044, Jiangsu, China Evgenios Agathokleous * Research Center for Global Changes and Ecosystem Carbon Sequestration & Mitigation, School of Ecology and Applied
Meteorology, Nanjing University of Information Science & Technology, Nanjing, 210044, Jiangsu, China Evgenios Agathokleous * Department of Environmental Health Sciences, Morrill I, N344,
University of Massachusetts, Amherst, MA, 01003, USA Edward J. Calabrese * Department of Agricultural Sciences, Biotechnology and Food Science, Cyprus University of Technology, Lemesos,
Cyprus Vasileios Fotopoulos Authors * Evgenios Agathokleous View author publications You can also search for this author inPubMed Google Scholar * Edward J. Calabrese View author
publications You can also search for this author inPubMed Google Scholar * Vasileios Fotopoulos View author publications You can also search for this author inPubMed Google Scholar
CONTRIBUTIONS E.A. conceived the manuscript and wrote the first draft. E.J.C. and V.F. revised the manuscript. E.A., E.J.C. and V.F. approved the final version for submission and eventual
publication. CORRESPONDING AUTHOR Correspondence to Evgenios Agathokleous. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION
PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. RIGHTS AND PERMISSIONS OPEN ACCESS This article is
licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any
medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed
material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are
included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and
your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this
licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Agathokleous, E., Calabrese, E.J. & Fotopoulos, V.
Low-dose stress promotes sustainable food production. _npj Sustain. Agric._ 2, 19 (2024). https://doi.org/10.1038/s44264-024-00026-0 Download citation * Received: 06 May 2024 * Accepted: 16
September 2024 * Published: 01 October 2024 * DOI: https://doi.org/10.1038/s44264-024-00026-0 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content:
Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative