Play all audios:
ABSTRACT Major depressive disorder (MDD) is a heterogeneous illness for which there are currently no effective methods to objectively assess severity, endophenotypes, or response to
treatment. Increasing evidence suggests that circulating levels of peripheral/serum growth factors and cytokines are altered in patients with MDD, and that antidepressant treatments reverse
or normalize these effects. Furthermore, there is a large body of literature demonstrating that MDD is associated with changes in endocrine and metabolic factors. Here we provide a brief
overview of the evidence that peripheral growth factors, pro-inflammatory cytokines, endocrine factors, and metabolic markers contribute to the pathophysiology of MDD and antidepressant
response. Recent preclinical studies demonstrating that peripheral growth factors and cytokines influence brain function and behavior are also discussed along with their implications for
diagnosing and treating patients with MDD. Together, these studies highlight the need to develop a biomarker panel for depression that aims to profile diverse peripheral factors that
together provide a biological signature of MDD subtypes as well as treatment response. SIMILAR CONTENT BEING VIEWED BY OTHERS SERUM BDNF LEVELS CORRELATE WITH REGIONAL CORTICAL THICKNESS IN
MINOR DEPRESSION: A PILOT STUDY Article Open access 03 September 2020 FIBROBLAST GROWTH FACTOR 21 (FGF21) IS INCREASED IN MDD AND INTERACTS WITH BODY MASS INDEX (BMI) TO AFFECT DEPRESSION
TRAJECTORY Article Open access 11 January 2022 RETINOID HOMEOSTASIS IN MAJOR DEPRESSIVE DISORDER Article Open access 23 February 2023 INTRODUCTION Major depressive disorder (MDD) is a
prevalent, heterogeneous illness characterized by depressed mood, anhedonia, and altered cognitive function. The lifetime prevalence of MDD is approximately 17% of the population and results
in tremendous secondary costs to society (Kessler et al, 2005; Wang et al, 2003). Diagnosis and treatment of MDD is based on relatively subjective assessments of diverse symptoms
representing multiple endophenotypes. To date, the biological bases for the heterogeneity of MDD remain poorly defined. Toward this goal, identification of biological markers could improve
the diagnosis and classification of MDD subtypes, as well as stratify patients into more homogeneous, clinically distinct subpopulations. Despite decades of searching, a non-invasive,
quantitative clinical test to aid in the diagnosis and treatment of MDD remains elusive (Lakhan et al, 2010). However, recent studies of MDD provide renewed hope. While there is no clear
single biomarker, there is mounting evidence of multiple dysregulated contributing factors, including growth factors and/or pro-inflammatory cytokines (Castren and Rantamaki, 2010; Krishnan
and Nestler, 2008; Miller et al, 2009; Schmidt and Duman, 2007). In addition, there is a long history and clear evidence for altered endocrine factors (eg, hypothalamic–pituitary–adrenal
(HPA), thyroid, sex steroids) and metabolic dysregulation (eg, insulin resistance) in mood disorders (Hendrickx et al, 2005). Thus, a viable alternative to the single-biomarker approach
could be the development of biomarker panels that aim to profile a diverse array of peripheral/serum growth factors, cytokines, hormones, and metabolic markers, to provide coverage of
multiple biological abnormalities that contribute to the heterogeneity of MDD, as well as treatment response. This endeavor will require a large number of patient samples to define severity,
subgroups, and response, but analytical tools are currently available to make biomarker assessment possible. In this review, we provide a brief overview of the key growth factors,
cytokines, hormones, and metabolic markers that could be included in an initial multi-analyte biomarker panel of MDD. Providing further support for this approach are recent preclinical
studies demonstrating that serum growth factors and cytokines can influence brain function, including cellular and behavioral responses. These findings indicate that analyses of these
factors will not only serve a biomarker function, but will also provide information about the underlying neurobiology of MDD subtypes, which will allow improved diagnosis and individualized
treatments. WHAT IS A BIOMARKER? The term biomarker can be used in a variety of ways. The most common biomarker concepts include specific features of an individual that are useful in
distinguishing the presence or absence of a disease state (‘diagnostic biomarkers’), or that predict treatment response (‘treatment biomarkers’). With regard to treatment biomarkers, the
biomarker may either be present at baseline and predict response to treatment, or, alternatively, may change in the short term in such a way as to predict the ultimate response. In the
latter case, the biomarker would be measured at baseline and again early in the course of therapy; ideally, lack of change in the marker would lead to alteration of the course of treatment.
In general, biomarkers are measurable features of an individual that represent indicators of a disease state or outcome with treatment. Moreover, biomarkers are typically thought of as a
biological feature (eg, genome variation, plasma concentration of a protein, etc), but do not have to be limited in this manner (Perlis, 2011). Most biomarkers are discovered initially in a
type of retrospective analysis of existing data sets. This, for example, was how a variety of gene variants were found to be associated with antidepressant treatment outcome in the Sequenced
Treatment Alternatives to Relieve Depression (STAR*D) study (Laje et al, 2009). In this case, as in others, the specific genetic variants were assayed in a _post-hoc_ manner, demonstrating
some degree of factor loading with response. However, alternative prospective designs can be employed by using a type of enrichment strategy. In an enriched design, biomarkers may be used to
select people into a clinical trial to maximize response to a given intervention, particularly enhancing drug–placebo differences. Biomarker designs, then, may be used to minimize sample
size to test for a therapeutic effect. A similar strategy is the ‘biomarker stratified design,’ in which there is a randomization in order to balance the distribution of a particular marker
(Perlis, 2011). This approach can be used to actually test for the differential usefulness of a biomarker in predicting differential responsiveness to a treatment. In the case of treatment
response, analysis of biomarkers represents a variation of mediator and moderator analyses as proposed by Baron and Kenny (1986). As elaborated by Kraemer et al (2002b), treatment moderators
are factors that ‘specify for whom or under what conditions the treatment works … They also suggest to clinicians which of their patients might be most responsive to the treatment and for
which patients other, more appropriate, treatments might be sought.’ Treatment biomarkers can serve as a special case of a biomarker that ‘labels’ the likelihood of responding to a given
treatment. A positive moderator, then, indicates the selection of a particular treatment and a negative moderator suggests choosing an alternative. A prescriptive moderator would favor one
treatment against another. Again, as stated by Kraemer et al (2002b), ‘moderators may also provide unique new and valuable information to guide future restructuring of diagnostic
classification and treatment decision making.’ A number of pharmacogenomic studies have evaluated the moderating effect of specific genetic variation on response to antidepressant therapies.
For example, as summarized recently by Lin and Chen (2008), the STAR*D study found single-nucleotide polymorphisms (SNPs) in several genes associated with response or adverse effects with
the SSRI antidepressant citalopram, subsequent antidepressants, or combinations of treatments. These included FK506-binding protein-5 (_FKBP5_), glutamate receptor ionotropic kainate-1
(_GRIK1_) and 4 (_GRIK4_), n-methyl-d-aspartate receptor-2A (_GRIN2A_), 5-hydroxytryptamine receptor-2A (_HTR2A_), potassium channel subfamily-K member-2 (_KCNK2_) (six SNPs), and the
serotonin transporter (_SLC6A4_) long/short variants. Several genes were also associated with treatment-emergent suicidality, including, cyclic-AMP response element-binding protein-1
(_CREB1_), glutamate receptor ionotropic AMPA-3 (_GRIA3_), and _GRIK2_. Other biological factors have been shown to be associated with lesser response to antidepressant therapy (Papakostas
and Fava, 2008). For example, greater number or size of white matter hyper-intensities on structural magnetic resonance imaging brain scans (presumably indicating small vessel disease) are
associated with reduced response to antidepressants (Alexopoulos et al, 2008; Iosifescu et al, 2006; Papakostas et al, 2005). Higher baseline levels of anxiety and overall depression
severity have been shown to be predictors of poorer response to antidepressant therapy (Papakostas and Fava, 2008). Fournier et al (2009) recently reported examples of clinical prescriptive
moderators of antidepressant response _vs_ cognitive behavioral psychotherapy (CBT) treatment of depression. In the original study, CBT and the antidepressant paroxetine were equally helpful
and more effective than placebo in a large MDD sample (DeRubeis et al, 2005). However, a secondary moderator analysis found that chronic depression, older age, and lower intelligence each
predicted relatively poor response across both treatments. Three prescriptive variables, being married, unemployment, and having experienced a greater number of recent stressful life events,
were identified and each predicted superior response to cognitive therapy relative to antidepressant medications. A mediator, on the other hand, is a factor that changes along with response
to a particular intervention (Kraemer et al, 2002b). In many cases, a mediator is a fundamental causal mechanism by which a particular treatment produces a change, but this does not have to
be the case. There can be special cases of secondary effects of treatments that are affected by the treatment and are associated with response to treatment but may not be the actual causal
mechanism that produces improvement on an illness measure. However, ideally, the mediator variable will change in such a way as to indicate subsequent improvement before the change in the
underlying disease state is manifest. In addition, the absence of early change in the mediator should predict lack of subsequent improvement in the disease-specific variable (eg, symptom
measure). Therefore, the mediator variable is most useful as an early marker of subsequent improvement in the disease state, and, therefore, would be a practical guide to treatment
prediction. A lack of early change in the mediator variable would indicate the need to change the treatment. The personality trait of neuroticism has been examined as a mediator variable for
response to SSRI treatment (Quilty et al, 2008). Neuroticism is considered a personality trait (thought by some as an endophenotype), which is characterized by a tendency to experience
negative emotions such as anxiety, sadness, embarrassment, anger, guilt, or disgust in face of perceived or anticipated stressors. Therefore, neuroticism is a vulnerability factor to both
anxiety and depressive disorders. Quilty et al (2008) evaluated two models of the relationship between neuroticism and response to antidepressant therapy, a ‘mediation’ model (ie, SSRI →
Neuroticism Change → Depression Change) and a ‘complication’ model (ie, SSRI → Depression Change → Neuroticism Change), by using a maximum likelihood of estimation approach. The ‘mediation’
model best fit the SSRI response data, indicating that overall neuroticism change is associated with change in depression severity. In the case of both moderators and mediators of treatment
effects, the slope of change in the underlying disease state is predicted by the baseline level of the marker in the case of moderators or the rate of change in the marker in the case of
mediators (Kraemer et al, 2002b). Mediators can serve as moderators and vice versa, but this does not have to be the case. Diagnostic biomarkers represent a different type of moderator
analysis. In this case, the presence of a marker indicates a higher likelihood of an underlying disease state and may be present before a disease is actually present (indicating a ‘risk’ or
‘vulnerability’ marker). As with treatment biomarkers, a diagnostic marker may or may not be directly related to the underlying causal mechanism for the condition. However, putative
biomarkers must be distinguished from associated features of a particular disease. For example, as noted below in the case of inflammatory factors such as elevations in cytokines, a
difference of a particular marker in a disease population in contrast to an unaffected group could be the result of an associated condition. In the case of inflammatory biomarkers, the
presence of an elevated cytokine may be a marker for depression, but it also could be associated with other conditions such as obesity, Type-2 diabetes, or cardiovascular disease that are
commonly associated with depression (Shelton and Miller, 2010). Therefore, matching of affected and unaffected groups should take co-varying features into consideration. Another inherent
limitation of biomarker identification has to do with the diagnostic accuracy of the typical clinical procedure used for identifying the disease state. In some cases, such as the
prostate-specific antigen (PSA) test for prostate cancer, the disease itself can be identified with high accuracy by prostate biopsy (Balk et al, 2003) (although, notably, even in this
instance, the benefit of the PSA test for identifying prostate cancer has been called into question (Andriole et al, 2009)). However, a test is unlikely to be better than whatever method was
used to identify the population at risk; in the case of psychiatric disorders, identification of the affected state is usually through a diagnostic interview. Hence, in psychiatry, any
biomarker is not likely to be better at identifying the condition than the clinical interview used to diagnose people in a study. Therefore, a diagnostic biomarker test is predominantly
useful in situations in which an extensive clinical interview is not feasible, such as in large-scale screenings. Alternatively, a biomarker discovered to be useful in accurately
distinguishing affected and unaffected people might be present in the case of people not yet affected and be a risk marker for the disease state before it is actually present. Although some
biomarkers are truly dichotomous, as in the case of the presence of a specific SNP or the repeat region of a gene, many others are actually continuous variables (eg, the plasma concentration
of a specific protein). In this case, a criterion cutoff will have to eventually be specified (Kraemer et al, 2002a). This process is consistent with a receiver operating characteristic
(ROC) analysis, in which different criterion levels can be used to create binary outcomes (eg, presence _vs_ absence of a condition or response _vs_ non-response to a treatment) based on a
classifier—in this case, a biomarker (Kraemer et al, 2002a). The ROC expresses the sensitivity (ie, true positive rate _vs_ the rate of false positives) and the specificity (true negatives
_vs_ false negatives) of a specific criterion level. Ideally, the criterion level will reflect a high degree of sensitivity and specificity of the classification threshold, again with
classification indicating either the presence (_vs_ absence) of the condition or the response (_vs_ non-response) to a treatment. For typical psychiatric diagnoses such as major depression
or schizophrenia, the likelihood of any given biomarker achieving a high enough degree of sensitivity and specificity—that is, an ideal ROC curve—to make the biomarker clinically useful is
relatively low. We propose that the use of multiple biomarkers may provide a possible solution to this problem. Although individual biomarkers may provide some greater level of true _vs_
false positive and negatives, the predictive abilities may improve when several different biomarkers are aggregated into a group, or biopanel, of predictor characteristics. Rather than
depending on a high level of predictive power of an individual marker, the biopanel approach would depend on an aggregate score or predictive algorithm for classification. Individual items
could then be added or subtracted to identify the best-performing set of predictor characteristics. In addition, the assessment of a panel of markers could potentially aid in the subdivision
of a heterogeneous illness that presents with a similar phenotype in a clinical interview. It is possible that individual biomarkers will aggregate in ways to inform the parsing of the MDD
phenotype into subtypes that may relate more closely to specific etiological pathways. Inflammatory cytokines and related factors, discussed in greater detail below, appear to more
consistently aggregate in individual patients but not in others. This type of clustering is likely to reflect something more closely related to an etiology of a subset of MDD. This, in turn,
could lead to more effective, etiologically based therapies for subgroups of patients. We will review a proposed set of biomarkers that should be considered for inclusion in future
biomarker studies, with a focus on growth factors, cytokines, and metabolic factors. GROWTH FACTORS A large body of evidence indicates that stress impairs trophic support whereas
antidepressants function, in part, to enhance trophic factor expression and neuroplasticity (Schmidt et al, 2008; Schmidt and Duman, 2007). Clinical studies demonstrate that patients with
MDD have altered blood/serum levels of growth factors. Consistent with these results, increasing evidence indicates that chronic stress exposure, which can precipitate or exacerbate
depressive episodes, alters the expression of growth factors, and that antidepressant treatment produces opposing effects. The following sections will discuss several of these key growth
factors, and will focus on (1) preclinical studies of stress and antidepressant regulation, and (2) clinical studies of blood of MDD patients. Evidence that peripheral administration of
these factors influences neuronal plasticity and behavior will also be discussed. BRAIN-DERIVED NEUROTROPHIC FACTOR Brain-derived neurotrophic factor (BDNF) regulates synaptic plasticity in
neuronal networks involved in depressive behaviors (Pittenger and Duman, 2007; Schinder and Poo, 2000). Regulation of BDNF may reverse stress-induced deficits in structural and synaptic
plasticity in the adult brain, resulting in cognitive flexibility and, subsequently, an increased ability to adapt/cope with environmental challenges that may precipitate or exacerbate
depressive episodes. Recent studies demonstrate that BDNF levels are decreased in the blood of MDD patients and reversed with antidepressant treatment (Brunoni et al, 2008b; Sen et al,
2008). INFLUENCE OF STRESS AND ANTIDEPRESSANTS ON BDNF Exposure to physical or psychological stressors leads to rapid downregulation of BDNF expression in the hippocampus, which could
contribute to experience-dependent modifications in neural networks that contribute to the pathogenesis of MDD (Nibuya et al, 1995, 1999; Rasmusson et al, 2002; Russo-Neustadt et al, 2001;
Smith et al, 1995b). By contrast, chronic antidepressant administration increases BDNF expression in the hippocampus (Altar et al, 2004; Newton et al, 2003; Nibuya et al, 1995;
Russo-Neustadt et al, 1999). Furthermore, recent studies have demonstrated that BDNF (ICV or intra-hippocampal) produces antidepressant behavioral responses in animal models of depression
(Hoshaw et al, 2005; Shirayama et al, 2002; Siuciak et al, 1997). Consistent with these findings, transgenic mice expressing a variant BDNF allele (Val66Met), which decreases the processing
and release of BDNF, are more vulnerable to stress-induced behavioral deficits and have an attenuated antidepressant response (Chen et al, 2006; Egan et al, 2003). BDNF deletion mutants also
show a depressive phenotype when exposed to mild stress (Duman et al, 2007), although there is no difference in behavior under non-stressed conditions (Chen et al, 2006; Monteggia et al,
2004; Saarelainen et al, 2003). Interestingly, clinical studies have reported a similar increase in stress vulnerability in subjects carrying the BDNF Val66Met polymorphism (Gatt et al,
2009). Postmortem studies report that hippocampal BDNF is decreased in MDD suicide subjects, but increased in subjects receiving antidepressant medication at the time of death (Chen et al,
2001b; Dwivedi et al, 2003; Karege et al, 2005). While there is compelling evidence that BDNF mediates the actions of antidepressants in the hippocampus, recent studies indicate that
increased BDNF/TrkB signaling has pro-depressive effects in other brain nuclei. For example, increased BDNF expression in the ventral tegmental area (VTA) promotes depressive-like behaviors
(Eisch et al, 2003). Consistent with these results, decreased VTA and nucleus accumbens BDNF produces antidepressant responses in a social defeat paradigm (Berton et al, 2006; Krishnan et
al, 2007b). Furthermore, overexpression of a dominant-negative form of TrkB in the nucleus accumbens results in an antidepressant response indicating that increased BDNF signaling has a
pro-depressive function in the ventral striatum (Eisch et al, 2003). Collectively these data indicate that the behavioral effects of BDNF and TrkB in animal models of depression are
region-specific, and that the pathogenesis of MDD is likely to include deficits in multiple brain regions. For these reasons, studies demonstrating antidepressant-like phenotypes in mutant
mice overexpressing BDNF or in mice receiving infusions of BDNF into the lateral ventricle may more accurately model the neuropathology of MDD than animal studies examining the role of BDNF
in one discrete brain region. Taken together, these studies indicate that reduced BDNF contributes to depressive behaviors in animal models and in humans, and that antidepressant treatment
increases or reverses these behavioral deficits by increasing BDNF. These findings are consistent with the hypothesis that the actions of antidepressants are due, in part, to BDNF-induced
neuronal plasticity and/or protection (Pittenger and Duman, 2007). BLOOD BDNF LEVELS ARE DECREASED IN PATIENTS WITH MDD A large number of clinical studies have reported that BDNF levels in
serum (Aydemir et al, 2006; Gervasoni et al, 2005; Karege et al, 2002; Shimizu et al, 2003) and plasma (Kim et al, 2007a; Lee et al, 2006) are significantly decreased in depressed patients,
and that this decrease is normalized by antidepressant treatments (Aydemir et al, 2005; Bocchio-Chiavetto et al, 2006; Gervasoni et al, 2005; Gonul et al, 2005; Huang et al, 2008; Okamoto et
al, 2008; Yoshimura et al, 2007; Zanardini et al, 2006), and confirmed by meta-analysis (Brunoni et al, 2008a; Sen et al, 2008). These findings indicate that blood BDNF may be a useful
biomarker, and that blood BDNF could have functional significance in the pathophysiology and/or treatment of mood disorders. BDNF is transcribed at relatively high levels and expressed in
peripheral tissues, including lung, heart, skeletal muscle, spleen, kidney, and blood (Braun et al, 1999; Koliatsos et al, 1993; Lommatzsch et al, 1999, 2005b; Nassenstein et al, 2003;
Scarisbrick et al, 1993; Timmusk et al, 1993; Yamamoto et al, 1996). Although the functional significance of these peripheral sources of BDNF is unknown, it is likely that BDNF in blood is
derived from these tissues as well as from brain, and that peripheral sources of BDNF contribute to reductions of blood BDNF in MDD patients. FUNCTIONAL SIGNIFICANCE PERIPHERAL BDNF The
possibility that peripheral growth factors can enter the brain and produce both behavioral and cellular responses is supported by studies of IGF-1 (Aberg et al, 2000; Duman et al, 2008b),
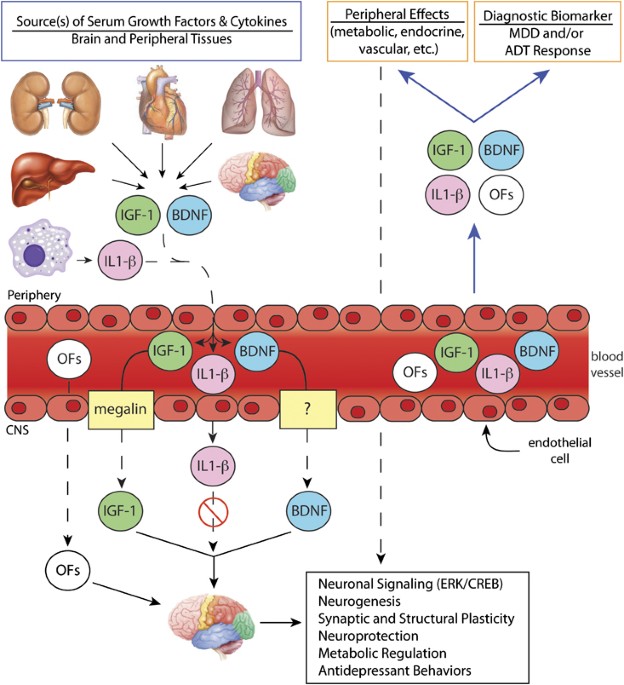
vascular endothelial growth factor (VEGF) (Fabel et al, 2003), and more recently of BDNF (Schmidt and Duman, 2010) (Figure 1). Chronic peripheral BDNF administration produces
antidepressant-like behavioral responses in animal models (Schmidt and Duman, 2010), effects that are similar to the actions of different classes of chemical antidepressants (Cryan et al,
2005), and partially blocks the effects of chronic unpredictable stress (Schmidt and Duman, 2010). These behavioral actions are associated with antidepressant cellular responses, including
increased survival of newborn hippocampal neurons (Schmidt and Duman, 2010), consistent with previous studies of central BDNF infusion (Sairanen et al, 2005). Peripheral BDNF administration
is also associated with increased BDNF levels in the brain and activation of the downstream signaling markers ERK and CREB (Schmidt and Duman, 2010). Despite studies demonstrating a
pro-depressive effect of BDNF/CREB in the striatum (Berton et al, 2006; Eisch et al, 2003; Wallace et al, 2009), these results indicate that antidepressant effects predominate in response to
peripheral BDNF (Schmidt et al, 2008). While the peripheral source(s) of endogenous BDNF and mechanisms for transport into the brain are currently unknown, blood BDNF provides a novel
window into brain structure and function that is relevant to MDD (Schmidt and Duman, 2010). INSULIN-LIKE GROWTH FACTOR-1 Insulin-like growth factor-1 (IGF-1) regulates cell growth and
metabolism in peripheral tissues (Stewart and Rotwein, 1996). Although IGF-1 is produced primarily by the liver and circulates in the bloodstream, it is also produced in the central nervous
system, where it has a critical role in nerve growth and differentiation, as well as neurotransmitter synthesis and release (Anlar et al, 1999; Bondy and Lee, 1993; D’Ercole et al, 1996;
Werther et al, 1990). Interestingly, the adult brain contains high levels of IGF-1 receptors, but unlike the developing brain, the expression levels of IGF-1 are low, suggesting that the
adult brain may utilize IGF-1 from peripheral sources (Bondy and Lee, 1993). INFLUENCE OF STRESS AND ANTIDEPRESSANTS ON IGF-1 Chronic antidepressant administration increases IGF-1 expression
in the rat brain (Khawaja et al, 2004), and IGF-1 regulates adult hippocampal neurogenesis (Anderson et al, 2002) and produces antidepressant behavioral responses (Hoshaw et al, 2005;
Malberg et al, 2007). Moreover, IGF signaling is altered in postmortem brain tissue in subjects with bipolar disorder (Bezchlibnyk et al, 2007). These results suggest that IGF-1 could
contribute to the cellular and behavioral responses to antidepressant treatments, as well as the pathophysiology of mood disorders. Peripheral IGF-1 crosses the blood–brain barrier through a
transporter-mediated mechanism (Carro et al, 2005; Pan and Kastin, 2000) and influences neuronal function (Pulford and Ishii, 2001; Reinhardt and Bondy, 1994). Physical exercise stimulates
the expression and release of liver IGF-1, and results in elevated brain uptake (Carro et al, 2000). Peripheral IGF-1 administration increases hippocampal neurogenesis (Aberg et al, 2000),
and blockade of peripheral IGF-1 reduces exercise-induced neurogenesis (Duman et al, 2008a; Trejo et al, 2001). These findings indicate that peripheral IGF-1 is transported into the brain,
where it produces cellular and behavioral responses. BLOOD IGF-1 LEVELS IN PATIENTS WITH ALTERED MOOD There have not been sufficient studies to determine whether peripheral IGF-1 is altered
in depressed patients or following antidepressant administration. However, exercise is associated with improved mood and increased serum IGF-1 expression in naïve elderly subjects (Cassilhas
et al, 2010). Additional studies are needed to determine the role of IGF-1 in MDD and response to antidepressants. FUNCTIONAL SIGNIFICANCE OF PERIPHERAL IGF-1 Peripheral IGF-1
administration has been shown to alter behavior independent of exercise and produce antidepressant-like behavioral responses. Peripheral IGF-1 administration reduces immobility in the FST
(Duman et al, 2009), and produces antidepressant behavioral responses in mice exposed to chronic unpredictable stress (Duman et al, 2009). Elevated blood IGF-1 levels are also associated
with increased adult hippocampal neurogenesis, improved cognition, and some of the beneficial effects of exercise, including reduced anxiety (Trejo et al, 2008, 2007). The behavioral effects
of peripherally administered IGF-1 are associated with increased levels of exogenous IGF-1 in the brain (Duman et al, 2009). Although speculative, some cases of MDD could result from
dysfunction of the peripheral expression and/or the transport of IGF-1 into the brain. VASCULAR ENDOTHELIAL GROWTH FACTOR VEGF is an endothelial cell mitogen and survival factor that
regulates vascular function (Leung et al, 1989), but is also expressed in the brain and has neuroprotective and neurogenic effects (Jin et al, 2002; Storkebaum et al, 2004; Warner-Schmidt
and Duman, 2007). INFLUENCE OF STRESS AND ANTIDEPRESSANTS ON VEGF Chronic stress exposure has been shown to decrease (Heine et al, 2005) and antidepressant administration to increase
hippocampal VEGF (Altar et al, 2004; Warner-Schmidt and Duman, 2007). Furthermore, impaired VEGF signaling in the brain blocks the effects of chemical antidepressants (Warner-Schmidt and
Duman, 2007) and exercise (Fabel et al, 2003) on hippocampal neurogenesis. Pharmacological antagonism of VEGF-mediated signaling in the brain blocks the behavioral effects of antidepressants
in rodent models (Greene et al, 2009; Lee et al, 2009; Warner-Schmidt and Duman, 2007). Peripheral VEGF also has a critical role in the neurogenic effects of exercise, which demonstrates
that blood VEGF has functional effects in the brain (Fabel et al, 2003). Taken together, these results indicate that VEGF is necessary and sufficient for the neurogenic and behavioral
actions of antidepressants. BLOOD VEGF LEVELS IN PATIENTS WITH ALTERED MOOD Clinical studies of peripheral VEGF in MDD are mixed. One study reports that VEGF expression is increased in
peripheral leukocytes of patients with MDD and that antidepressant treatment reverses these effects (Iga et al, 2006). Consistent with these results, blood VEGF levels are increased in
patients with MDD (Kahl et al, 2009). By contrast, another study found no significant differences in blood VEGF levels between patients with MDD and healthy controls, and following
antidepressant treatment (Ventriglia et al, 2009). Moreover, preclinical findings indicate that serum VEGF levels are not different in a genetic rat model of depression (Elfving et al,
2010). These divergent clinical findings are likely due to significant differences in patient populations, including age, gender, total number of depressive episodes (ie, recurrent _vs_
acute), and comorbid disorders. However, these clinical findings suggest that blood VEGF levels may be differentially altered depending upon the endophenotype of MDD studied, but further
studies are needed and warranted. OTHER GROWTH FACTORS Other growth factors that may also serve as biomarkers of MDD and/or antidepressant response include glial cell line-derived
neurotrophic factor (GDNF) and fibroblast growth factor-2 (FGF-2), both of which are altered in humans with MDD (Kahl et al, 2009; Rosa et al, 2006; Takebayashi et al, 2006). FGF-2, FGF
receptors, and the GDNF receptor are altered by antidepressant treatment (Chen et al, 2001a; Evans et al, 2004; Gaughran et al, 2006). FGF-2, neurotrophin-3 (NT-3), and nerve growth factor
(NGF) influence adult hippocampal neurogenesis and/or are regulated by stress and antidepressant treatments, and could contribute to stress-induced cellular and behavioral deficits, and
antidepressant responses (Dwivedi et al, 2005; Hock et al, 2000; Lu et al, 2005; Mallei et al, 2002; Molteni et al, 2001; Smith et al, 1995a). Finally, peripheral VGF expression is decreased
in patients with MDD (Cattaneo et al, 2010) and administration of recombinant VGF produces antidepressant behavioral responses in mice (Hunsberger et al, 2007). While the exact role of
GDNF, FGF-2, NT-3, NGF, and VGF in the pathogenesis of MDD and/or antidepressant behavioral and cellular responses is unclear, there is sufficient evidence to implicate these growth factors
in MDD. CYTOKINES AND INFLAMMATORY MARKERS Increasing evidence indicates that inflammation may have a critical role in the pathophysiology of MDD (Miller et al, 2009). Clinical studies
demonstrate that patients with MDD have elevated blood/serum levels of inflammatory markers, including pro-inflammatory cytokines. Consistent with these results, inhibiting pro-inflammatory
cytokine signaling in patients with inflammatory disorders, as well as patients with MDD, improves mood and facilitates antidepressant treatment response. Furthermore, chronic stress
exposure alters the expression of cytokines, and antidepressant treatment neutralizes these effects. The following sections will discuss preclinical and clinical studies of several of these
key cytokines in rodent models and MDD patients. TNF-_Α_ AND IL-6 MDD is also accompanied by altered immune function and activation of the inflammatory response system (Dinan, 2009).
Activated macrophages secrete pro-inflammatory cytokines, which may contribute to MDD. Cytokine activation produces sickness behaviors, which share features with depression (Dunn et al,
2005; Koo and Duman, 2008). Moreover, chronic stress exposure produces changes in immune function that may influence the pathophysiology of MDD (Miller et al, 2009). CELLULAR AND BEHAVIORAL
ACTIONS OF TNF-_Α_ AND IL-6 The pro-inflammatory cytokines interleukin-6 (IL-6) and tumor necrosis factor-_α_ (TNF-_α_) have direct inhibitory effects on adult hippocampal neurogenesis
(Iosif et al, 2006; Monje et al, 2003), and, therefore, may attenuate antidepressant efficacy by decreasing hippocampal neurogenesis or interfering with the neurogenic properties of
antidepressants. In addition, mutant mice lacking TNF-_α_ receptors show antidepressant behavioral phenotypes (Simen et al, 2006). Taken together, these preclinical findings suggest that
TNF-_α_ and IL-6 may block the behavioral and cellular responses to antidepressants and/or facilitate depressive phenotypes. BLOOD TNF-_Α_ AND IL-6 IN PATIENTS WITH ALTERED MOOD
Peripheral/serum levels of IL-6 and TNF-_α_ are increased in patients with MDD (Dowlati et al, 2010; Kahl et al, 2006), and these effects are normalized following antidepressant treatment
(Levine et al, 1999; Raison et al, 2006; Sluzewska et al, 1996; Steptoe et al, 2007; Thomas et al, 2005; Tuglu et al, 2003). These consistent clinical findings indicate that TNF-_α_ and IL-6
are putative biomarkers of depressive episodes and treatment response. Interestingly, treatment-resistant patients with MDD have elevated blood IL-6 levels compared with
treatment-responsive patients (Maes et al, 1997). Therefore, changes in blood IL-6 levels may serve as a marker to track those patient populations that respond to a given antidepressant
treatment. FUNCTIONAL SIGNIFICANCE OF TNF-_Α_ AND IL-6 Preclinical studies of cytokines and depressive behaviors correlate with clinical studies of depression (Khairova et al, 2009). For
example, immunotherapy using IL-2 or interferon-_α_ (IFN-_α_) is associated with cognitive impairments and depressed mood that correlate with elevated blood levels of IFN-_α_, IL-6, IL-8,
and IL-10 (Bonaccorso et al, 2002, 2001; Capuron et al, 2001a, 2001b; Dieperink et al, 2000). Depression, anxiety, and memory impairments are also associated with immune activation by the
bacterial endotoxin LPS in healthy subjects and are associated with increased blood IL-1 and TNF-_α_ (Yirmiya et al, 2000). Increasing evidence suggests that patients with MDD have an
imbalance between pro- and anti-inflammatory cytokines that can be normalized following antidepressant treatment (Kim et al, 2007b; Sutcigil et al, 2007; Taler et al, 2007). Some patients
with MDD also have abnormal allelic variants of the genes for IL-1 and TNF-_α_, and those with elevated levels of TNF-_α_ have an attenuated therapeutic response to antidepressant treatment
(Eller et al, 2008; Fertuzinhos et al, 2004; Khairova et al, 2009). Clinical studies also demonstrate that cytokine antagonists have antidepressant behavioral effects, even in the absence of
an immune challenge. The TNF-_α_ antagonists etanercept and infliximab attenuate depressive symptoms induced by immune activation during psoriasis (Krishnan et al, 2007a; Tyring et al,
2006). There is also a report that the cyclooxygenase-2 (COX2) inhibitor celexocib, which inhibits the production of pro-inflammatory cytokines, including TNF-_α_ and IL-1_β_, produces a
rapid antidepressant response in patients with MDD (Muller et al, 2006). Taken together, these findings raise the possibility that reductions in inflammatory processes might contribute to
treatment response, and that inhibiting pro-inflammatory signaling may be a promising strategy to treat depressed patients with increased blood cytokine profiles. INTERLEUKIN-1_Β_ EFFECTS OF
STRESS AND ANTIDEPRESSANTS ON IL-1_Β_ Recent evidence indicates that dysregulation of pro-inflammatory cytokines, including IL-1_β_, influences the etiology and/or pathophysiology of MDD
(Raison et al, 2006). Elevated levels of pro-inflammatory cytokines may also contribute to the damaging effects of stress. Stress exposure increases IL-1_β_ in the hippocampus (Johnson et
al, 2005; Nguyen et al, 1998), IL-1_β_ inhibits adult hippocampal neurogenesis, and blockade of IL-1 inhibits the effects of stress on neurogenesis (Koo and Duman, 2008). Increased IL-1_β_
in the hippocampus is also associated with stress-induced impairments in synaptic plasticity (Murray and Lynch, 1998) as well as activation of the HPA axis (Linthorst et al, 1994; Rivier,
1993; Sapolsky et al, 1987). Administration of an IL-1_β_ receptor antagonist into the hippocampus blocks the BDNF decrease caused by stress, suggesting that the anti-neurogenic effects of
cytokines may be mediated, in part, through regulation of BDNF (Barrientos et al, 2003) and/or IGF-1 (O’Connor et al, 2008). Thus, stress-induced deficits and hippocampal plasticity are
regulated by complex mechanisms involving cytokines and growth factors. BLOOD IL-1_Β_ LEVELS IN PATIENTS WITH ALTERED MOOD Blood IL-1_β_ is increased in patients with MDD (Diniz et al, 2010;
Thomas et al, 2005) and antidepressant treatment may reverse this effect (Himmerich et al, 2010; Song et al, 2009). However, not all clinical studies demonstrate increased circulating
levels of IL-1_β_ in patients with MDD (Jazayeri et al, 2010) and these changes are not as consistent as those observed when examining IL-6, TNF-_α_, and C-reactive protein (CRP) (Howren et
al, 2009b). These mixed clinical results are likely due to heterogeneity of MDD. FUNCTIONAL SIGNIFICANCE OF PERIPHERAL IL-1_Β_ Peripheral and central IL-1_β_ administration induces sickness
behaviors, including anorexia, weight loss, anhedonia, fatigue, impaired social interaction, and memory dysfunction, symptoms that are also observed in patients with MDD (Goshen and Yirmiya,
2009; Koo and Duman, 2008). By contrast, inhibition of IL-1_β_ signaling blocks depressive and sickness-related behaviors (Goshen and Yirmiya, 2009; Koo and Duman, 2008). Recent studies
demonstrate that impaired IL-1 receptor signaling blocks stress-induced anhedonia (Goshen and Yirmiya, 2008; Koo and Duman, 2008) and produces antidepressant effects in an animal model of
behavioral despair (Maier and Watkins, 1995). Future studies are required to identify the precise mechanism(s) by which peripheral/serum IL-1_β_ activates HPA function and produces anhedonic
and anxiogenic behavioral responses. OTHER CYTOKINES/INFLAMMATORY MARKERS The risk of developing MDD is increased in patients undergoing cytokine or IFN therapy for the treatment of cancer
or viral infection such as hepatitis-C (Capuron and Dantzer, 2003). A recent study demonstrated that IFN therapy-induced depressive episodes are associated with decreased blood BDNF levels,
suggesting a point of intersection with stress and antidepressant treatments (Kenis et al, 2010). These results indicate that cytokines and IFNs significantly contribute to the effects of
stress, as well as the precipitation and maintenance of MDD, and conversely that neutralization of these cytokines could have antidepressant effects (Dantzer et al, 2008). Additional work is
needed to determine the role of other cytokines, including IL-4, IL-2, IL-8, IL-10, and/or IFN-_γ_, in MDD. High-sensitivity CRP (hs-CRP), a marker of low-grade inflammation, is a
cardiovascular disease risk factor and a potential biomarker of immunological activation (De Berardis et al, 2006). Coronary artery disease is associated with a high incidence of MDD
(Nemeroff et al, 1998) and with higher levels of circulating hs-CRP (Pearson et al, 2003), which is synthesized in the liver in response to stimulation from IL-6 and IL-1. A meta-analysis
reveals positive associations between MDD and hs-CRP, IL-6, and, to a lesser extent, IL-1 (Howren et al, 2009a). These findings highlight a role for hs-CRP and its precursors as
mediator/moderator factors of depression, although its precise role remains unclear. Mood disorders could also result from acquired immune disorders. Prolonged activation of the peripheral
immune system as occurs during systemic infections, cancer, or autoimmune disorders results in immune signaling in the brain that can lead to the development of depressive episodes (Dantzer
et al, 2008). Recent findings indicate that soluble IL-2 receptor levels (a marker of T-cell activation) are increased in patients with MDD (Mossner et al, 2007). Collectively these results
suggest that both acquired (eg, T- and B-cell) and innate (eg, macrophage) immune response may have critical roles in the pathophysiology of MDD. However, it remains unclear whether
activation of inflammatory signaling during depression is an indirect result of peripheral processes and/or whether stress exposure induces inflammatory responses directly within the brain
(Miller et al, 2009). DIRECT _VS_ INDIRECT EFFECTS OF PERIPHERAL FACTORS ON NEURONAL FUNCTION It remains to be determined whether the behavioral and cellular actions of peripheral BDNF, as
well as other growth factors and cytokines, are mediated by direct actions on the brain and/or indirect mechanisms through regulation of peripheral endocrine or metabolic actions. There are
reports that peripheral BDNF can cross the blood–brain barrier, possibly through active transport similar to IGF-1 (Carro et al, 2005; Trejo et al, 2007), although this remains controversial
(Pan et al, 1998; Pardridge, 2002; Poduslo and Curran, 1996). In addition, saturable transport systems from blood to the brain have been described for cytokines including IL-1_β_, IL-6, and
TNF-_α_ (Banks, 2005). Therefore, circulating BDNF and other growth factors may be transported into the brain and have direct effects on neuronal as well as glial function. While much is
known about the roles of peripheral IGF-1 in metabolic processes and peripheral cytokines in inflammatory processes, the functional significance of blood BDNF derived from peripheral tissues
is unclear. Moreover, the mechanisms that regulate blood BDNF, IGF-1, and cytokines during MDD have not been identified. Future studies to identify the mechanisms (ie, transcriptional,
synthesis, release, clearance, etc) underlying the regulation of peripheral as well as central expression of growth factors and cytokines will further elucidate the neurobiology of mood
disorders. An often-overlooked question with regard to putative biomarkers is the relationship between peripheral and central changes in biomarker levels. It is not clear whether altered
levels of putative biomarkers in peripheral tissues must mirror changes in the brain and vice versa. Future studies directly addressing this question will aid in classifying biomarkers as
moderators, mediators, diagnostic markers, or a combination of these roles. ENDOCRINE AND METABOLIC MARKERS Analyses of stress-induced changes of peripheral endocrine and metabolic markers
will also aid in the diagnosis and treatment of MDD. An extensive literature now demonstrates that neuroendocrine and metabolic functions are altered in patients with MDD. NEUROENDOCRINE
FUNCTION AND MDD Depression is associated with altered regulation of the HPA axis that results in increased release of corticotropin-releasing hormone (CRH) and in some cases sustained
elevation of cortisol (Nestler et al, 2002). Glucocorticoids (cortisol in humans and corticosterone in rodents) bind to their receptors in the HPA axis and act as negative regulators of HPA
axis activity. Increased activity of the HPA axis in MDD is due, in part, to altered feedback inhibition of the HPA axis by endogenous glucocorticoids (for further review see, Pariante,
2009). Impaired negative feedback of the HPA axis by glucocorticoids is mediated, in part, by altered expression of the glucocorticoid receptor (Pariante and Miller, 2001). It has been
proposed that elevated cortisol in patients with MDD is a compensatory mechanism in response to decreased glucocorticoid receptor function and expression in the brain (Raison and Miller,
2003). Preclinical studies demonstrate that chronic antidepressant administration leads to the upregulation of glucocorticoid receptor expression and function, and thus increased negative
feedback regulation of the HPA axis (Pariante and Miller, 2001). Biomarker panels that monitor changes in cortisol, as well as other HPA axis factors (eg, CRF), will provide important
information for characterization of MDD subtypes. Cortisol, however, is not elevated in all persons with MDD. Some data indicate that persons with the melancholic subtype of MDD may be more
likely to have increased HPA axis activity than non-melancholic patients (Gold and Chrousos, 2002; Wong et al, 2000). Melancholia is a distinct form of depression characterized by
consistently down and non-reactive mood, anhedonia, decreased sleep and appetite, and weight loss (Fink and Taylor, 2007). Persons with melancholia are more likely to have elevations in
plasma cortisol and lack of dexamethasone suppression relative to non-melancholic patients (Gold and Chrousos, 2002), which tend to normalize with effective treatment (Fink and Taylor,
2007). Inflammatory markers, including cytokines, regulate neuroendocrine function. Acute cytokine administration is associated with increased expression and release of CRH,
adrenocorticotropic hormone (ACTH), and cortisol (Besedovsky and del Rey, 1996). Cytokines may impair neuroendocrine function by interfering with the negative feedback regulation of the HPA
axis, a hallmark of MDD that is reflected by decreased responsiveness to glucocorticoids (Miller et al, 2009). Increased cytokine signaling inhibits glucocorticoid receptor function and
increases the expression of the relatively inert _β_-isoform, while decreasing the expression of the active _α_-isoform, of the glucocorticoid receptor (Pace et al, 2007). In addition,
glucocorticoids have clear inhibitory effects on inflammation (Rhen and Cidlowski, 2005). Dysregulation of the exquisite balance between HPA axis sensitivity to glucocorticoids and the
innate immune system (Miller et al, 2009) can be readily monitored in MDD patients. Therefore, biomarker panels of MDD should target pathways by which the immune system impacts the brain,
including cytokines, inflammatory mediators (eg, COX-2, prostaglandin), reactive nitrogen and oxygen species (eg, nitric oxide, hydrogen peroxide), monoamines, neurotrophic factors, and HPA
axis hormones (eg, CRH, cortisol) and receptors (eg, glucocorticoid receptors). Monitoring these putative biomarkers during antidepressant treatment may aid in identifying patient
populations that are responsive to inflammation-targeted therapies (Miller et al, 2009). METABOLIC FUNCTION AND MDD Circulating hormones such as leptin and ghrelin relay information
pertaining to peripheral energy homeostatic levels to the brain (Lutter and Nestler, 2009). Low levels of leptin have been found to be associated with depressive behaviors in humans and
rodents (Lu, 2007), and chronic stress exposure decreases serum leptin (Lu et al, 2006). Consistent with these results, acute leptin administration produces antidepressant responses (Liu et
al, 2010) and leptin administration blocks depressive behavior in leptin-deficient mice, effects that are associated with increased hippocampal BDNF expression (Yamada et al, 2011). By
contrast, chronic stress exposure increases serum ghrelin levels (Lutter et al, 2008). Calorie restriction produces antidepressant responses that are associated with increased circulating
ghrelin levels (Lutter et al, 2008). Collectively, these results suggest that ghrelin counteracts stress-induced behavioral deficits by promoting antidepressant responses. Thus, leptin and
ghrelin may serve as putative biomarkers for MDD in general or in depressed patients with altered metabolic function. Metabolic syndrome is a cluster of cardiovascular risk factors that are
associated with increased incidence of cardiovascular disease and diabetes. Metabolic syndrome is also associated with MDD (Skilton et al, 2007). Antidepressants exert variable effects on
the constituent components of metabolic syndrome (McIntyre et al, 2010). A recent study suggests that decreased HDL cholesterol levels, but not other markers of metabolic syndrome, may
predict the development of new-onset MDD in pre-elderly populations (Akbaraly et al, 2011). This finding is consistent with the hypothesis that dyslipidemia mediates depressive episodes in
the elderly (Ancelin et al, 2010). While future studies are required to determine the exact role of dyslipidemia in the etiology of MDD, HDL levels may predict the onset of an MDD
endophenotype that manifests later in life. Depression is frequently associated with comorbid disorders, including diabetes, a metabolic disorder that is associated with the damaging effects
of inflammation and oxidative stress in the brain (Hendrickx et al, 2005). Type-2 diabetes is characterized by hyperglycemia and the inability of the body to control blood glucose levels.
Type-2 diabetes usually begins as insulin resistance, a disorder in which glucose uptake by peripheral cells is impaired, which leads to a compensatory increase in insulin secretion by the
pancreas. Eventually, the pancreas can no longer produce enough insulin to maintain euglycemia and Type-2 diabetes occurs. However, the relationship between MDD and insulin resistance is not
clear (Adriaanse et al, 2006; Lawlor et al, 2003; Pan et al, 2008; Qiuhua et al, 2011; Timonen et al, 2005, 2006). These studies varied significantly in patient demographics, gender,
depression ratings, and insulin resistance measurements. Diabetes-induced elevations in blood glucose and insulin levels produce inflammatory effects in the brain and may contribute to the
development of MDD (Hendrickx et al, 2005). Therefore, a biomarker panel of MDD should track insulin resistance and glucose levels as potential mediators of MDD in pre-diabetic and diabetic
patients, respectively. Changes in serum lipid profiles and free radicals should also be considered as future studies determine the extent of these changes in metabolic disorders and the
concurrence of depressive episodes. Further evidence for a role of metabolic dysregulation is provided by studies demonstrating that impaired peripheral glucose regulation is associated with
cognitive decline and depression, especially in obese subjects and patients with Type-2 diabetes (Hendrickx et al, 2005). The negative consequences of aberrant glycemic control on brain
function are mediated, in part, by insulin, glucose, growth factors, cortisol, cytokines, and reactive oxygen species (Hendrickx et al, 2005). Specifically, diabetes and metabolic syndrome
are associated with increased HPA axis activity, and some of the factors that regulate diabetes-related cognitive decline include peripheral IGF-1 and cortisol (Hendrickx et al, 2005). Thus,
the etiology and pathophysiology of MDD appear to be tightly regulated by complex interplays between endocrine, immune, and metabolic systems. Although there is not a clear understanding of
how these systems function together to mediate depressive episodes, biomarker panels that monitor these peripheral factors will provide descriptive evidence toward this goal. NON-PROTEOMIC
BIOMARKERS OF MDD Genetic factors have a critical role in the development of MDD and provide insights into the mechanisms underlying depression. Candidate gene studies have implicated
polymorphisms in the genes encoding the serotonin transporter, serotonin receptor-2A, BDNF, and tryptophan hydroxylase in MDD (Lohoff, 2010). These studies along with genome-wide association
studies have not identified a single common gene variant that increases the risk of MDD substantially (Lohoff, 2010). Instead, depression is likely to result from complex interactions
between multiple genetic and environmental factors. Thus, tracking genetic variants in patient blood may serve to compliment biomarker panels by providing more information relating genotype
to MDD and treatment response. An emerging literature indicates that stress exposure induces epigenetic mechanisms such as histone modifications and DNA methylation that promote maladaptive
behaviors. Chronic social stress decreases hippocampal BDNF through long-lasting dimethlyation of histones at the level of BDNF promoters and is associated with a pro-depressive phenotype
(Tsankova et al, 2006). By contrast, chronic antidepressant administration reverses stress-induced BDNF repression through epigenetic mechanisms involving histone-3 acetylation and histone-3
lysine-4 methylation (Tsankova et al, 2006). Moreover, systemic administration of a DNA methylation inhibitor produces antidepressant behavioral responses that are associated with decreased
DNA methylation and increased BDNF expression in the hippocampus (Sales et al, 2011). Stressful events in early life also produce long-lasting epigenetic marks that influence affect and
mood. Offspring of mothers with low levels of nurturing behavior had increased methylation of the glucocorticoid receptor variant GR17 promoter, which leads to decreased GR17 expression in
adulthood (Weaver et al, 2004). Thus, long-lasting epigenetic modifications have a critical role in stress-induced and antidepressant behavioral responses. However, these studies to date
have focused on the transcriptional regulation of BDNF and glucocorticoid receptor genes in the hippocampus. It remains to be determined whether epigenetic changes in response to stress or
antidepressant treatment can be monitored from components of blood and cerebral spinal fluid to aid in the diagnosis of MDD. In addition to comprehensive proteomic screens, future biomarkers
of MDD and antidepressant response are likely to include epigenetic and genetic factors. Recently, more comprehensive approaches to identifying diagnostic biomarkers of mood disorders
including MDD have been described. Convergent Functional Genomics is a multidisciplinary method that integrates animal model gene expression data with human genetic linkage/association data,
as well as human tissue (ie, postmortem brain, blood, etc) data, to identify and prioritize candidate genes and molecular substrates for subsequent hypothesis-driven research. Using gene
arrays to examine blood biomarker genes, Convergent Functional Genomics has identified genes associated specifically with high or low mood states (Le-Niculescu et al, 2009). These results
are consistent with previous studies demonstrating differential expression of these genes in postmortem brain tissue from mood disorder subjects (Le-Niculescu et al, 2009). Identifying
genetic and proteomic biomarkers for psychiatric disorders including MDD is limited by cost, lack of predictability, and unreliability due to polygenetic inheritance and environmental
influences (Lakhan et al, 2010). It remains to be determined whether any of the genetic biomarker panels identified using Convergent Functional Genetics and other techniques correlate with
treatment response and whether these methods could be used to differentiate MDD severity and/or subtypes. SPECIFICITY OF BIOMARKERS FOR MOOD DISORDERS Altered blood levels of BDNF, IGF-1,
and cytokines are not specific to MDD. Peripheral BDNF and IGF-1 levels are decreased in several psychiatric illnesses, including eating disorders (Nakazato et al, 2003; Saito et al, 2009),
schizophrenia (Green et al, 2010; Toyooka et al, 2002), and/or panic (Kobayashi et al, 2005). Furthermore, there is a high incidence of comorbid or coincident diseases, including Type-2
diabetes and MDD (Katon, 2008), as well as strong associations between MDD and metabolic syndrome (Dunbar et al, 2008). Alterations of serum growth factors and cytokines have also been
demonstrated in cardiovascular (Ejiri et al, 2005; Kaplan et al, 2005; von der Thusen et al, 2003), inflammatory (Katsanos et al, 2001; Lee et al, 2010; Lommatzsch et al, 2005a;
Schulte-Herbruggen et al, 2005), and metabolic diseases (Dunger et al, 2003; Han et al, 2010; Kaldunski et al, 2010), all of which are more common in depressed patients than the general
population (Shelton and Miller, 2010). However, patients with these conditions but without depression (ie, persons with cardiovascular disease or Type-2 diabetes) will have altered levels of
the putative biomarkers described above. These findings suggest that altered peripheral systems contribute to a broader disease state. Monitoring multiple factors will provide a more
complete assessment and thereby identify a spectrum of factors that better characterize disease state as well as specific disease symptoms. This information can also be used for targeted
treatment to augment or neutralize altered growth factor or cytokine levels. Stated simply, whereas single biomarkers are unlikely to adequately distinguish depressed from non-depressed
subjects, panels of multiple biomarkers may work significantly better. Biomarker panels for simultaneous detection of peripheral cytokines, growth factors, hormones, and other protein
markers will allow the identification of a peripheral signature that differentiates MDD subtypes and distinguishes MDD from other disorders (Figure 2). Identifying proteomic biomarkers for
psychiatric disorders will require a large sample size in order to demonstrate that these methods are both predictable and reliable. Furthermore, it will be necessary to demonstrate that
biomarker panels correlate with antidepressant efficacy, severity, and/or endophenotypes of MDD in independent cohorts of patients. Nevertheless, the information provided by such panels will
be invaluable for showing imbalances of multiple systems and will aid in the treatment and management of illness. Clinical studies examining putative biomarkers have compared changes in
patients with MDD _vs_ matched, healthy control subjects. Therefore, an important limitation to biomarker selection for depression is the lack of direct comparisons between MDD and other
disorders with comorbid depression. Identifying novel candidate biological markers for MDD by using proteomic profiling methods will enable simultaneous detection of cytokines, growth
factors, hormones, and other protein markers in plasma samples in order to determine a peripheral signature for MDD. Furthermore, multi-analyte biomarker panels may provide differential
diagnoses between MDD and other disorders with depression as a symptom. However, more clinical studies directly comparing changes in peripheral biomarkers in MDD compared with other comorbid
disorders are warranted to determine whether changes in putative biomarkers are specific to MDD. Interestingly, a recent study demonstrated that plasma biomarker profiling has the potential
to differentiate psychiatric disorders by identifying unique biomarkers for each condition (Domenici et al, 2010). Thus, comprehensive biomarker panels provide peripheral signatures that
differentiate psychiatric disorders and may facilitate differential diagnoses of MDD subtypes and comorbid disorders. Predictive algorithms may be successfully derived from multiple
biological variables of MDD. Algorithms that predict inclusion in depressed and control groups can be thought of as a set of partial differential equations that aim to achieve maximum
separation between these two defined groups. Individual biomarkers of a predictor set might be unlikely to provide sufficient predictive power to produce adequate separation between groups.
However, group identity may be optimized by using equation modeling in which each individual variable has a designated weight, depending on its positive predictive value (PPV) (Altman and
Bland, 1994). While the use of multiple biomarkers for diagnosis and treatment prediction is not generally done in psychiatry, it is a common strategy in other fields such as oncology (Dunn
et al, 2011; Malinowski, 2007a, 2007b; Marrero et al, 2010; Vauthey et al, 2010; Yurkovetsky et al, 2010; Zhu et al, 2011). A simple clinical example would be methods for staging of various
cancers that may use multiple predictive characteristics (Vauthey et al, 2010). It remains unclear, however, as to how comorbid psychiatric disorders, including substance use disorders, will
affect prediction algorithms for the diagnosis of MDD. PPV (also known as precision rate) is the proportion of tested individuals with positive test results who are correctly diagnosed
(Altman and Bland, 1994). In order for a diagnostic test to be clinically relevant, it will have to have a PPV that is robust to comorbidity. While a test might be developed in a
non-comorbid group, it would ultimately have to be tested in samples specifically selected for high rates of medical and psychiatric comorbidity. CONCLUSIONS Clinical and preclinical studies
have identified a number of factors that may serve as putative biomarkers for diagnosing and treating MDD. However, the utility of any given growth factor, cytokine, endocrine factor, or
metabolic marker to serve as a clinically useful biomarker of MDD is limited by a lack of sensitivity and specificity. Therefore, we propose a panel of multiple biomarkers to improve the
predictive power of these factors as measured using an aggregate score or predictive algorithm to diagnose and classify MDD subtypes as well as measure treatment response. A number of
questions regarding peripheral/blood biomarkers and MDD remain. First, the optimal time point at which peripheral/blood biomarkers should be measured during the day and during treatment is
not clear. There are also potential confounds in interpreting changes in biomarkers during antidepressant treatment. For example, it remains uncertain whether clear distinctions in biomarker
levels will differentiate antidepressant efficacy or remission. Finally, it is not clear whether putative biomarkers for MDD have sufficient sensitivity, specificity, and reproducibility
for predicting therapeutic responses and remission rates that are reliable to diagnose and treat patients with MDD (Leuchter et al, 2010). One approach that could address these issues is the
use of a stress, immune, and/or metabolic challenge test in MDD, to reveal altered regulation of peripheral biomarkers. This would be analogous to a stress test used for cardiovascular
disease or glucose tolerance/insulin resistance for diabetes. By comparing pre- and post-test levels of blood biomarkers, this type of challenge could reveal more robust abnormalities in the
regulation of growth factors, cytokines, endocrine, and metabolic markers. Challenge paradigms are routinely used for other medical conditions and could provide an important approach for
the diagnosis and treatment of mood disorders. Developing an operational biomarker panel of MDD will require significant effort and resources. Successful implementation of a biomarker panel
capable of tracking endophenotype signatures and treatment response must provide comprehensive coverage of multiple biological systems. While putative biomarkers of MDD have been identified,
further studies are needed to classify these factors as mediators, moderators, or diagnostic markers. Large network collaborations will be key to obtaining sufficient power as large sample
sizes will be essential to define severity and parse MDD into identifiable subtypes. Results obtained from biomarker panels will need to be standardized such that clear associations between
these signatures and current clinical definitions of heterogenous subtypes of MDD are readily apparent. In line with these goals, operational definitions of MDD as set forth by the DSM-V and
future terminology must recognize and classify depression as multiple disorders. Significant financial resources and sustained investigation will be required as initial biomarker panels are
updated and better-performing measures are introduced. As new biomarkers are identified, it is likely that multiple panels will be needed to diagnosis MDD, monitor disease
progression/severity, and select an appropriate treatment. While acute or chronic stressors may induce depressive episodes in some individuals, most people are resilient to these effects.
Therefore, it is conceivable that markers of stress resilience may be identified. Recent studies have begun to investigate the biological bases underlying stress resilience with the hope of
identifying protective factors that may promote resilience in individuals who cannot successfully adapt to stress (Feder et al, 2009). Critical individual differences in resilience to the
behavioral and neurochemical effects of stress have been reported (Feder et al, 2009). For example, a recent study demonstrated that increased hippocampal BDNF mediates resilience in rodents
exposed to chronic stress (Taliaz et al, 2011). Moreover, peripheral BDNF administration partially attenuates stress-induced behavioral deficits (Schmidt and Duman, 2010). Taken together,
these results suggest that BDNF may serve as a putative resilience marker. Resilience is regulated by neuroadaptations in neural circuits that regulate fear (Bush et al, 2007), reward (Cao
et al, 2010), social behavior (Elliott et al, 2010), affect, and mood (Wager et al, 2008). Future studies are required to identify molecular substrates that may serve as resilience
biomarkers and pharmacotherapeutic targets to promote resilient phenotypes. In summary, a growing body of evidence indicates that MDD is associated with decreased expression of
peripheral/serum growth factors as well as increased levels of circulating cytokines. Antidepressant treatment normalizes or reverses many of these effects. In addition, recent evidence
indicates that peripheral/serum BDNF and IGF-1 produce antidepressant effects in behavioral and cellular models of depression, and that blocking IL-1_β_- and TNF-_α_-mediated signaling
attenuates stress-induced behavioral and cellular deficits in rodents and humans. Therefore, peripheral/serum BDNF, IGF-1, and cytokines may serve not only as biomarkers of MDD and treatment
response, but also have functional consequences. The heterogeneity of MDD and concurrent changes in the expression of these peripheral proteins in comorbid psychiatric, immune,
inflammatory, and metabolic disorders renders the selection of one individual biomarker for MDD outdated and impractical. Instead, new methods that simultaneously profile a diversity of
peripheral biomarkers will inform the diagnosis of MDD, including heterogeneous subtypes and the response to antidepressant treatments. REFERENCES * Aberg MA, Aberg ND, Hedbacker H,
Oscarsson J, Eriksson PS (2000). Peripheral infusion of IGF-I selectively induces neurogenesis in the adult rat hippocampus. _J Neurosci_ 20: 2896–2903. Article CAS PubMed PubMed Central
Google Scholar * Adriaanse MC, Dekker JM, Nijpels G, Heine RJ, Snoek FJ, Pouwer F (2006). Associations between depressive symptoms and insulin resistance: the Hoorn Study. _Diabetologia_
49: 2874–2877. Article CAS PubMed Google Scholar * Akbaraly TN, Ancelin ML, Jaussent I, Ritchie C, Barberger-Gateau P, Dufouil C _et al_ (2011). Metabolic syndrome and onset of
depressive symptoms in the elderly: findings from the three-city study. _Diabetes Care_ 34: 904–909. Article PubMed PubMed Central Google Scholar * Alexopoulos GS, Murphy CF,
Gunning-Dixon FM, Latoussakis V, Kanellopoulos D, Klimstra S _et al_ (2008). Microstructural white matter abnormalities and remission of geriatric depression. _Am J Psychiatry_ 165: 238–244.
Article PubMed Google Scholar * Altar CA, Laeng P, Jurata LW, Brockman JA, Lemire A, Bullard J _et al_ (2004). Electroconvulsive seizures regulate gene expression of distinct
neurotrophic signaling pathways. _J Neurosci_ 24: 2667–2677. Article CAS PubMed PubMed Central Google Scholar * Altman DG, Bland JM (1994). Diagnostic tests 2: predictive values. _BMJ_
309: 102. Article CAS PubMed PubMed Central Google Scholar * Ancelin ML, Carriere I, Boulenger JP, Malafosse A, Stewart R, Cristol JP _et al_ (2010). Gender and genotype modulation of
the association between lipid levels and depressive symptomatology in community-dwelling elderly (the ESPRIT study). _Biol Psychiatry_ 68: 125–132. Article CAS PubMed Google Scholar *
Anderson MF, Aberg MA, Nilsson M, Eriksson PS (2002). Insulin-like growth factor-I and neurogenesis in the adult mammalian brain. _Brain Res Dev Brain Res_ 134: 115–122. Article CAS PubMed
Google Scholar * Andriole GL, Crawford ED, Grubb III RL, Buys SS, Chia D, Church TR _et al_ (2009). Mortality results from a randomized prostate-cancer screening trial. _N Engl J Med_
360: 1310–1319. Article CAS PubMed PubMed Central Google Scholar * Anlar B, Sullivan KA, Feldman EL (1999). Insulin-like growth factor-I and central nervous system development. _Horm
Metab Res_ 31: 120–125. Article CAS PubMed Google Scholar * Aydemir C, Yalcin ES, Aksaray S, Kisa C, Yildirim SG, Uzbay T _et al_ (2006). Brain-derived neurotrophic factor (BDNF) changes
in the serum of depressed women. _Prog Neuropsychopharmacol Biol Psychiatry_ 30: 1256–1260. Article CAS PubMed Google Scholar * Aydemir O, Deveci A, Taneli F (2005). The effect of
chronic antidepressant treatment on serum brain-derived neurotrophic factor levels in depressed patients: a preliminary study. _Prog Neuropsychopharmacol Biol Psychiatry_ 29: 261–265.
Article CAS PubMed Google Scholar * Balk SP, Ko YJ, Bubley GJ (2003). Biology of prostate-specific antigen. _J Clin Oncol_ 21: 383–391. Article CAS PubMed Google Scholar * Banks WA
(2005). Blood-brain barrier transport of cytokines: a mechanism for neuropathology. _Curr Pharm Des_ 11: 973–984. Article CAS PubMed Google Scholar * Baron RM, Kenny DA (1986). The
moderator-mediator variable distinction in social psychological research: conceptual, strategic, and statistical considerations. _J Pers Soc Psychol_ 51: 1173–1182. Article CAS PubMed
Google Scholar * Barrientos RM, Sprunger DB, Campeau S, Higgins EA, Watkins LR, Rudy JW _et al_ (2003). Brain-derived neurotrophic factor mRNA downregulation produced by social isolation is
blocked by intrahippocampal interleukin-1 receptor antagonist. _Neuroscience_ 121: 847–853. Article CAS PubMed Google Scholar * Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W,
Russo SJ _et al_ (2006). Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. _Science_ 311: 864–868. Article CAS PubMed Google Scholar * Besedovsky HO, del
Rey A (1996). Immune-neuro-endocrine interactions: facts and hypotheses. _Endocr Rev_ 17: 64–102. Article CAS PubMed Google Scholar * Bezchlibnyk YB, Xu L, Wang JF, Young LT (2007).
Decreased expression of insulin-like growth factor binding protein 2 in the prefrontal cortex of subjects with bipolar disorder and its regulation by lithium treatment. _Brain Res_ 1147:
213–217. Article CAS PubMed Google Scholar * Bocchio-Chiavetto L, Zanardini R, Bortolomasi M, Abate M, Segala M, Giacopuzzi M _et al_ (2006). Electroconvulsive therapy (ECT) increases
serum brain derived neurotrophic factor (BDNF) in drug resistant depressed patients. _Eur Neuropsychopharmacol_ 16: 620–624. Article CAS PubMed Google Scholar * Bonaccorso S, Marino V,
Biondi M, Grimaldi F, Ippoliti F, Maes M (2002). Depression induced by treatment with interferon-alpha in patients affected by hepatitis C virus. _J Affect Disord_ 72: 237–241. Article CAS
PubMed Google Scholar * Bonaccorso S, Puzella A, Marino V, Pasquini M, Biondi M, Artini M _et al_ (2001). Immunotherapy with interferon-alpha in patients affected by chronic hepatitis C
induces an intercorrelated stimulation of the cytokine network and an increase in depressive and anxiety symptoms. _Psychiatry Res_ 105: 45–55. Article CAS PubMed Google Scholar * Bondy
CA, Lee WH (1993). Patterns of insulin-like growth factor and IGF receptor gene expression in the brain. Functional implications. _Ann NY Acad Sci_ 692: 33–43. Article CAS PubMed Google
Scholar * Braun A, Lommatzsch M, Mannsfeldt A, Neuhaus-Steinmetz U, Fischer A, Schnoy N _et al_ (1999). Cellular sources of enhanced brain-derived neurotrophic factor production in a mouse
model of allergic inflammation. _Am J Respir Cell Mol Biol_ 21: 537–546. Article CAS PubMed Google Scholar * Brunoni AR, Lopes M, Fregni F (2008a). A systematic review and meta-analysis
of clinical studies on major depression and BDNF levels: implications for the role of neuroplasticity in depression. _Int J Neuropsychopharmacol_ 11: 1169–1180. Article CAS PubMed Google
Scholar * Brunoni AR, Lopes M, Fregni F (2008b). A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: implications for the role of neuroplasticity
in depression. _Int J Neuropsychopharmacol_ 11: 1169–1180. Article CAS PubMed Google Scholar * Bush DE, Sotres-Bayon F, LeDoux JE (2007). Individual differences in fear: isolating fear
reactivity and fear recovery phenotypes. _J Trauma Stress_ 20: 413–422. Article PubMed Google Scholar * Cao JL, Covington III HE, Friedman AK, Wilkinson MB, Walsh JJ, Cooper DC _et al_
(2010). Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. _J Neurosci_ 30: 16453–16458. Article CAS PubMed PubMed
Central Google Scholar * Capuron L, Dantzer R (2003). Cytokines and depression: the need for a new paradigm. _Brain Behav Immun_ 17 (Suppl 1): S119–S124. Article CAS PubMed Google
Scholar * Capuron L, Ravaud A, Dantzer R (2001a). Timing and specificity of the cognitive changes induced by interleukin-2 and interferon-alpha treatments in cancer patients. _Psychosom
Med_ 63: 376–386. Article CAS PubMed Google Scholar * Capuron L, Ravaud A, Gualde N, Bosmans E, Dantzer R, Maes M _et al_ (2001b). Association between immune activation and early
depressive symptoms in cancer patients treated with interleukin-2-based therapy. _Psychoneuroendocrinology_ 26: 797–808. Article CAS PubMed Google Scholar * Carro E, Nunez A, Busiguina
S, Torres-Aleman I (2000). Circulating insulin-like growth factor I mediates effects of exercise on the brain. _J Neurosci_ 20: 2926–2933. Article CAS PubMed PubMed Central Google
Scholar * Carro E, Spuch C, Trejo JL, Antequera D, Torres-Aleman I (2005). Choroid plexus megalin is involved in neuroprotection by serum insulin-like growth factor I. _J Neurosci_ 25:
10884–10893. Article CAS PubMed PubMed Central Google Scholar * Cassilhas RC, Antunes HK, Tufik S, de Mello MT (2010). Mood, anxiety, and serum IGF-1 in elderly men given 24 weeks of
high resistance exercise. _Percept Mot Skills_ 110: 265–276. Article PubMed Google Scholar * Castren E, Rantamaki T (2010). The role of BDNF and its receptors in depression and
antidepressant drug action: reactivation of developmental plasticity. _Dev Neurobiol_ 70: 289–297. Article CAS PubMed Google Scholar * Cattaneo A, Sesta A, Calabrese F, Nielsen G, Riva
MA, Gennarelli M (2010). The expression of VGF is reduced in leukocytes of depressed patients and it is restored by effective antidepressant treatment. _Neuropsychopharmacology_ 35:
1423–1428. Article CAS PubMed PubMed Central Google Scholar * Chen AC, Eisch AJ, Sakai N, Takahashi M, Nestler EJ, Duman RS (2001a). Regulation of GFRalpha-1 and GFRalpha-2 mRNAs in rat
brain by electroconvulsive seizure. _Synapse_ 39: 42–50. Article CAS PubMed Google Scholar * Chen B, Dowlatshahi D, MacQueen GM, Wang JF, Young LT (2001b). Increased hippocampal BDNF
immunoreactivity in subjects treated with antidepressant medication. _Biol Psychiatry_ 50: 260–265. Article CAS PubMed Google Scholar * Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao
CJ _et al_ (2006). Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. _Science_ 314: 140–143. Article CAS PubMed PubMed Central Google Scholar * Cryan JF,
Page ME, Lucki I (2005). Differential behavioral effects of the antidepressants reboxetine, fluoxetine, and moclobemide in a modified forced swim test following chronic treatment.
_Psychopharmacology (Berl)_ 182: 335–344. Article CAS Google Scholar * Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW (2008). From inflammation to sickness and depression: when
the immune system subjugates the brain. _Nat Rev Neurosci_ 9: 46–56. CAS PubMed PubMed Central Google Scholar * De Berardis D, Campanella D, Gambi F, La Rovere R, Carano A, Conti CM _et
al_ (2006). The role of C-reactive protein in mood disorders. _Int J Immunopathol Pharmacol_ 19: 721–725. Article CAS PubMed Google Scholar * D’Ercole AJ, Ye P, Calikoglu AS,
Gutierrez-Ospina G (1996). The role of the insulin-like growth factors in the central nervous system. _Mol Neurobiol_ 13: 227–255. Article PubMed Google Scholar * DeRubeis RJ, Hollon SD,
Amsterdam JD, Shelton RC, Young PR, Salomon RM _et al_ (2005). Cognitive therapy _vs_ medications in the treatment of moderate to severe depression. _Arch Gen Psychiatry_ 62: 409–416.
Article PubMed Google Scholar * Dieperink E, Willenbring M, Ho SB (2000). Neuropsychiatric symptoms associated with hepatitis C and interferon alpha: a review. _Am J Psychiatry_ 157:
867–876. Article CAS PubMed Google Scholar * Dinan TG (2009). Inflammatory markers in depression. _Curr Opin Psychiatry_ 22: 32–36. Article PubMed Google Scholar * Diniz BS, Teixeira
AL, Talib L, Gattaz WF, Forlenza OV (2010). Interleukin-1beta serum levels is increased in antidepressant-free elderly depressed patients. _Am J Geriatr Psychiatry_ 18: 172–176. Article
PubMed Google Scholar * Domenici E, Wille DR, Tozzi F, Prokopenko I, Miller S, McKeown A _et al_ (2010). Plasma protein biomarkers for depression and schizophrenia by multi analyte
profiling of case–control collections. _PLoS One_ 5: e9166. Article PubMed PubMed Central CAS Google Scholar * Dowlati Y, Herrmann N, Swardfager W, Liu H, Sham L, Reim EK _et al_
(2010). A meta-analysis of cytokines in major depression. _Biol Psychiatry_ 67: 446–457. Article CAS PubMed Google Scholar * Duman CH, Schlesinger L, Kodama M, Russell DS, Duman RS
(2007). A role for MAP kinase signaling in behavioral models of depression and antidepressant treatment. _Biol Psychiatry_ 61: 661–670. Article CAS PubMed Google Scholar * Duman CH,
Schlesinger L, Russell DS, Duman RS (2008a). Voluntary exercise produces antidepressant and anxiolytic behavioral effects in mice. _Brain Res_ 1199: 148–158. Article CAS PubMed PubMed
Central Google Scholar * Duman CH, Schlesinger L, Terwilliger R, Russell DS, Newton SS, Duman RS (2008b). Peripheral insulin-like growth factor-I produces antidepressant-like behavior and
contributes to the effect of exercise. _Behav Brain Res_ 198: 366–371. Article PubMed PubMed Central CAS Google Scholar * Duman CH, Schlesinger L, Terwilliger R, Russell DS, Newton SS,
Duman RS (2009). Peripheral insulin-like growth factor-I produces antidepressant-like behavior and contributes to the effect of exercise. _Behav Brain Res_ 198: 366–371. Article CAS PubMed
Google Scholar * Dunbar JA, Reddy P, Davis-Lameloise N, Philpot B, Laatikainen T, Kilkkinen A _et al_ (2008). Depression: an important comorbidity with metabolic syndrome in a general
population. _Diabetes Care_ 31: 2368–2373. Article PubMed PubMed Central Google Scholar * Dunger DB, Ong KK, Sandhu MS (2003). Serum insulin-like growth factor-I levels and potential
risk of type 2 diabetes. _Horm Res_ 60 (Suppl 3): 131–135. CAS PubMed Google Scholar * Dunn AJ, Swiergiel AH, de Beaurepaire R (2005). Cytokines as mediators of depression: what can we
learn from animal studies? _Neurosci Biobehav Rev_ 29: 891–909. Article CAS PubMed Google Scholar * Dunn BK, Jegalian K, Greenwald P (2011). Biomarkers for early detection and as
surrogate endpoints in cancer prevention trials: issues and opportunities. _Recent Results Cancer Res_ 188: 21–47. Article CAS PubMed Google Scholar * Dwivedi Y, Mondal AC, Rizavi HS,
Conley RR (2005). Suicide brain is associated with decreased expression of neurotrophins. _Biol Psychiatry_ 58: 315–324. Article CAS PubMed Google Scholar * Dwivedi Y, Rizavi HS, Conley
RR, Roberts RC, Tamminga CA, Pandey GN (2003). Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. _Arch Gen
Psychiatry_ 60: 804–815. Article CAS PubMed Google Scholar * Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A _et al_ (2003). The BDNF val66met polymorphism
affects activity-dependent secretion of BDNF and human memory and hippocampal function. _Cell_ 112: 257–269. Article CAS PubMed Google Scholar * Eisch AJ, Bolanos CA, de Wit J, Simonak
RD, Pudiak CM, Barrot M _et al_ (2003). Brain-derived neurotrophic factor in the ventral midbrain-nucleus accumbens pathway: a role in depression. _Biol Psychiatry_ 54: 994–1005. Article
CAS PubMed Google Scholar * Ejiri J, Inoue N, Kobayashi S, Shiraki R, Otsui K, Honjo T _et al_ (2005). Possible role of brain-derived neurotrophic factor in the pathogenesis of coronary
artery disease. _Circulation_ 112: 2114–2120. Article CAS PubMed Google Scholar * Elfving B, Plougmann PH, Wegener G (2010). Differential brain, but not serum VEGF levels in a genetic
rat model of depression. _Neurosci Lett_ 474: 13–16. Article CAS PubMed Google Scholar * Eller T, Vasar V, Shlik J, Maron E (2008). Pro-inflammatory cytokines and treatment response to
escitalopram in major depressive disorder. _Prog Neuropsychopharmacol Biol Psychiatry_ 32: 445–450. Article CAS PubMed Google Scholar * Elliott E, Ezra-Nevo G, Regev L, Neufeld-Cohen A,
Chen A (2010). Resilience to social stress coincides with functional DNA methylation of the Crf gene in adult mice. _Nat Neurosci_ 13: 1351–1353. Article CAS PubMed Google Scholar *
Evans SJ, Choudary PV, Neal CR, Li JZ, Vawter MP, Tomita H _et al_ (2004). Dysregulation of the fibroblast growth factor system in major depression. _Proc Natl Acad Sci USA_ 101:
15506–15511. Article CAS PubMed PubMed Central Google Scholar * Fabel K, Fabel K, Tam B, Kaufer D, Baiker A, Simmons N _et al_ (2003). VEGF is necessary for exercise-induced adult
hippocampal neurogenesis. _Eur J Neurosci_ 18: 2803–2812. Article PubMed Google Scholar * Feder A, Nestler EJ, Charney DS (2009). Psychobiology and molecular genetics of resilience. _Nat
Rev Neurosci_ 10: 446–457. Article CAS PubMed PubMed Central Google Scholar * Fertuzinhos SM, Oliveira JR, Nishimura AL, Pontual D, Carvalho DR, Sougey EB _et al_ (2004). Analysis of
IL-1alpha, IL-1beta, and IL-1RA [correction of IL-RA] polymorphisms in dysthymia. _J Mol Neurosci_ 22: 251–256. Article CAS PubMed Google Scholar * Fink M, Taylor MA (2007). Resurrecting
melancholia. _Acta Psychiatr Scand Suppl_ 433: 14–20. Article Google Scholar * Fournier JC, DeRubeis RJ, Shelton RC, Hollon SD, Amsterdam JD, Gallop R (2009). Prediction of response to
medication and cognitive therapy in the treatment of moderate to severe depression. _J Consult Clin Psychol_ 77: 775–787. Article PubMed PubMed Central Google Scholar * Gatt JM, Nemeroff
CB, Dobson-Stone C, Paul RH, Bryant RA, Schofield PR _et al_ (2009). Interactions between BDNF Val66Met polymorphism and early life stress predict brain and arousal pathways to syndromal
depression and anxiety. _Mol Psychiatry_ 14: 681–695. Article CAS PubMed Google Scholar * Gaughran F, Payne J, Sedgwick PM, Cotter D, Berry M (2006). Hippocampal FGF-2 and FGFR1 mRNA
expression in major depression, schizophrenia and bipolar disorder. _Brain Res Bull_ 70: 221–227. Article CAS PubMed Google Scholar * Gervasoni N, Aubry JM, Bondolfi G, Osiek C, Schwald
M, Bertschy G _et al_ (2005). Partial normalization of serum brain-derived neurotrophic factor in remitted patients after a major depressive episode. _Neuropsychobiology_ 51: 234–238.
Article CAS PubMed Google Scholar * Gold PW, Chrousos GP (2002). Organization of the stress system and its dysregulation in melancholic and atypical depression: high _vs_ low CRH/NE
states. _Mol Psychiatry_ 7: 254–275. Article CAS PubMed Google Scholar * Gonul AS, Akdeniz F, Taneli F, Donat O, Eker C, Vahip S (2005). Effect of treatment on serum brain-derived
neurotrophic factor levels in depressed patients. _Eur Arch Psychiatry Clin Neurosci_ 255: 381–386. Article PubMed Google Scholar * Goshen I, Kreisel T, Ben-Menachem-Zidon O, Licht T,
Weidenfeld J, Ben-Hur T _et al_ (2008). Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. _Mol
Psychiatry_ 13: 717–728. Article CAS PubMed Google Scholar * Goshen I, Yirmiya R (2009). Interleukin-1 (IL-1): a central regulator of stress responses. _Front Neuroendocrinol_ 30: 30–45.
Article CAS PubMed Google Scholar * Green MJ, Matheson SL, Shepherd A, Weickert CS, Carr VJ (2010). Brain-derived neurotrophic factor levels in schizophrenia: a systematic review with
meta-analysis. _Mol Psychiatry_. (e-pub ahead of print) * Greene J, Banasr M, Lee B, Warner-Schmidt J, Duman RS (2009). Vascular endothelial growth factor signaling is required for the
behavioral actions of antidepressant treatment: pharmacological and cellular characterization. _Neuropsychopharmacology_ 34: 2459–2468. Article CAS PubMed Google Scholar * Han JC,
Muehlbauer MJ, Cui HN, Newgard CB, Haqq AM (2010). Lower brain-derived neurotrophic factor in patients with Prader–Willi syndrome compared to obese and lean control subjects. _J Clin
Endocrinol Metab_ 95: 3532–3536. Article CAS PubMed PubMed Central Google Scholar * Heine VM, Zareno J, Maslam S, Joels M, Lucassen PJ (2005). Chronic stress in the adult dentate gyrus
reduces cell proliferation near the vasculature and VEGF and Flk-1 protein expression. _Eur J Neurosci_ 21: 1304–1314. Article PubMed Google Scholar * Hendrickx H, McEwen BS, Ouderaa F
(2005). Metabolism, mood and cognition in aging: the importance of lifestyle and dietary intervention. _Neurobiol Aging_ 26 (Suppl 1): 1–5. Article PubMed Google Scholar * Himmerich H,
Milenovic S, Fulda S, Plumakers B, Sheldrick AJ, Michel TM _et al_ (2010). Regulatory T cells increased while IL-1beta decreased during antidepressant therapy. _J Psychiatr Res_ 44:
1052–1057. Article PubMed Google Scholar * Hock C, Heese K, Muller-Spahn F, Huber P, Riesen W, Nitsch RM _et al_ (2000). Increased cerebrospinal fluid levels of neurotrophin 3 (NT-3) in
elderly patients with major depression. _Mol Psychiatry_ 5: 510–513. Article CAS PubMed Google Scholar * Hoshaw BA, Malberg JE, Lucki I (2005). Central administration of IGF-I and BDNF
leads to long-lasting antidepressant-like effects. _Brain Res_ 1037: 204–208. Article CAS PubMed Google Scholar * Howren MB, Lamkin DM, Suls J (2009a). Associations of depression with
C-reactive protein, IL-1, and IL-6: a meta-analysis. _Psychosom Med_ 71: 171–186. Article CAS PubMed Google Scholar * Howren MB, Lamkin DM, Suls J (2009b). Associations of depression
with C-reactive protein, IL-1, and IL-6: a meta-analysis. _Psychosom Med_ 71: 171–186. Article CAS PubMed Google Scholar * Huang TL, Lee CT, Liu YL (2008). Serum brain-derived
neurotrophic factor levels in patients with major depression: effects of antidepressants. _J Psychiatr Res_ 42: 521–525. Article PubMed Google Scholar * Hunsberger JG, Newton SS, Bennett
AH, Duman CH, Russell DS, Salton SR _et al_ (2007). Antidepressant actions of the exercise-regulated gene VGF. _Nat Med_ 13: 1476–1482. Article CAS PubMed Google Scholar * Iga JI, Ueno
SI, Yamauchi K, Numata S, Tayoshi-Shibuya S, Kinouchi S _et al_ (2006). Gene expression and association analysis of vascular endothelial growth factor in major depressive disorder. _Prog
Neuropsychopharmacol Biol Psychiatry_ 31: 658–663. Article PubMed CAS Google Scholar * Iosif RE, Ekdahl CT, Ahlenius H, Pronk CJ, Bonde S, Kokaia Z _et al_ (2006). Tumor necrosis factor
receptor 1 is a negative regulator of progenitor proliferation in adult hippocampal neurogenesis. _J Neurosci_ 26: 9703–9712. Article CAS PubMed PubMed Central Google Scholar *
Iosifescu DV, Renshaw PF, Lyoo IK, Lee HK, Perlis RH, Papakostas GI _et al_ (2006). Brain white-matter hyperintensities and treatment outcome in major depressive disorder. _Br J Psychiatry_
188: 180–185. Article PubMed Google Scholar * Jazayeri S, Keshavarz SA, Tehrani-Doost M, Djalali M, Hosseini M, Amini H _et al_ (2010). Effects of eicosapentaenoic acid and fluoxetine on
plasma cortisol, serum interleukin-1beta and interleukin-6 concentrations in patients with major depressive disorder. _Psychiatry Res_ 178: 112–115. Article CAS PubMed Google Scholar *
Jin K, Zhu Y, Sun Y, Mao XO, Xie L, Greenberg DA (2002). Vascular endothelial growth factor (VEGF) stimulates neurogenesis _in vitro_ and _in vivo_. _Proc Natl Acad Sci USA_ 99: 11946–11950.
Article CAS PubMed PubMed Central Google Scholar * Johnson JD, Campisi J, Sharkey CM, Kennedy SL, Nickerson M, Greenwood BN _et al_ (2005). Catecholamines mediate stress-induced
increases in peripheral and central inflammatory cytokines. _Neuroscience_ 135: 1295–1307. Article CAS PubMed Google Scholar * Kahl KG, Bens S, Ziegler K, Rudolf S, Dibbelt L, Kordon A
_et al_ (2006). Cortisol, the cortisol–dehydroepiandrosterone ratio, and pro-inflammatory cytokines in patients with current major depressive disorder comorbid with borderline personality
disorder. _Biol Psychiatry_ 59: 667–671. Article CAS PubMed Google Scholar * Kahl KG, Bens S, Ziegler K, Rudolf S, Kordon A, Dibbelt L _et al_ (2009). Angiogenic factors in patients with
current major depressive disorder comorbid with borderline personality disorder. _Psychoneuroendocrinology_ 34: 353–357. Article CAS PubMed Google Scholar * Kaldunski M, Jia S, Geoffrey
R, Basken J, Prosser S, Kansra S _et al_ (2010). Identification of a serum-induced transcriptional signature associated with type 1 diabetes in the BioBreeding rat. _Diabetes_ 59:
2375–2385. Article CAS PubMed PubMed Central Google Scholar * Kaplan RC, Strickler HD, Rohan TE, Muzumdar R, Brown DL (2005). Insulin-like growth factors and coronary heart disease.
_Cardiol Rev_ 13: 35–39. PubMed Google Scholar * Karege F, Perret G, Bondolfi G, Schwald M, Bertschy G, Aubry JM (2002). Decreased serum brain-derived neurotrophic factor levels in major
depressed patients. _Psychiatry Res_ 109: 143–148. Article CAS PubMed Google Scholar * Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R (2005). Neurotrophin levels in postmortem
brains of suicide victims and the effects of antemortem diagnosis and psychotropic drugs. _Brain Res Mol Brain Res_ 136: 29–37. Article CAS PubMed Google Scholar * Katon WJ (2008). The
comorbidity of diabetes mellitus and depression. _Am J Med_ 121 (11 Suppl 2): S8–15. Article PubMed PubMed Central Google Scholar * Katsanos KH, Tsatsoulis A, Christodoulou D, Challa A,
Katsaraki A, Tsianos EV (2001). Reduced serum insulin-like growth factor-1 (IGF-1) and IGF-binding protein-3 levels in adults with inflammatory bowel disease. _Growth Horm IGF Res_ 11:
364–367. Article CAS PubMed Google Scholar * Kenis G, Prickaerts J, van Os J, Koek GH, Robaeys G, Steinbusch HW _et al_ (2010). Depressive symptoms following interferon-alpha therapy:
mediated by immune-induced reductions in brain-derived neurotrophic factor? _Int J Neuropsychopharmacol_ 14: 247–253. Article PubMed CAS Google Scholar * Kessler RC, Chiu WT, Demler O,
Merikangas KR, Walters EE (2005). Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. _Arch Gen Psychiatry_ 62: 617–627.
Article PubMed PubMed Central Google Scholar * Khairova RA, Machado-Vieira R, Du J, Manji HK (2009). A potential role for pro-inflammatory cytokines in regulating synaptic plasticity in
major depressive disorder. _Int J Neuropsychopharmacol_ 12: 561–578. Article CAS PubMed Google Scholar * Khawaja X, Xu J, Liang JJ, Barrett JE (2004). Proteomic analysis of protein
changes developing in rat hippocampus after chronic antidepressant treatment: implications for depressive disorders and future therapies. _J Neurosci Res_ 75: 451–460. Article CAS PubMed
Google Scholar * Kim YK, Lee HP, Won SD, Park EY, Lee HY, Lee BH _et al_ (2007a). Low plasma BDNF is associated with suicidal behavior in major depression. _Prog Neuropsychopharmacol Biol
Psychiatry_ 31: 78–85. Article CAS PubMed Google Scholar * Kim YK, Na KS, Shin KH, Jung HY, Choi SH, Kim JB (2007b). Cytokine imbalance in the pathophysiology of major depressive
disorder. _Prog Neuropsychopharmacol Biol Psychiatry_ 31: 1044–1053. Article CAS PubMed Google Scholar * Kobayashi K, Shimizu E, Hashimoto K, Mitsumori M, Koike K, Okamura N _et al_
(2005). Serum brain-derived neurotrophic factor (BDNF) levels in patients with panic disorder: as a biological predictor of response to group cognitive behavioral therapy. _Prog
Neuropsychopharmacol Biol Psychiatry_ 29: 658–663. Article CAS PubMed Google Scholar * Koliatsos VE, Clatterbuck RE, Winslow JW, Cayouette MH, Price DL (1993). Evidence that
brain-derived neurotrophic factor is a trophic factor for motor neurons _in vivo_. _Neuron_ 10: 359–367. Article CAS PubMed Google Scholar * Koo JW, Duman RS (2008). IL-1beta is an
essential mediator of the antineurogenic and anhedonic effects of stress. _Proc Natl Acad Sci USA_ 105: 751–756. Article CAS PubMed PubMed Central Google Scholar * Kraemer HC, Schultz
SK, Arndt S (2002a). Biomarkers in psychiatry: methodological issues. _Am J Geriatr Psychiatry_ 10: 653–659. Article PubMed Google Scholar * Kraemer HC, Wilson GT, Fairburn CG, Agras WS
(2002b). Mediators and moderators of treatment effects in randomized clinical trials. _Arch Gen Psychiatry_ 59: 877–883. Article PubMed Google Scholar * Krishnan R, Cella D, Leonardi C,
Papp K, Gottlieb AB, Dunn M _et al_ (2007a). Effects of etanercept therapy on fatigue and symptoms of depression in subjects treated for moderate to severe plaque psoriasis for up to 96
weeks. _Br J Dermatol_ 157: 1275–1277. Article CAS PubMed Google Scholar * Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ _et al_ (2007b). Molecular adaptations underlying
susceptibility and resistance to social defeat in brain reward regions. _Cell_ 131: 391–404. Article CAS PubMed Google Scholar * Krishnan V, Nestler EJ (2008). The molecular neurobiology
of depression. _Nature_ 455: 894–902. Article CAS PubMed PubMed Central Google Scholar * Laje G, Perlis RH, Rush AJ, McMahon FJ (2009). Pharmacogenetics studies in STAR*D: strengths,
limitations, and results. _Psychiatr Serv_ 60: 1446–1457. Article PubMed PubMed Central Google Scholar * Lakhan SE, Vieira K, Hamlat E (2010). Biomarkers in psychiatry: drawbacks and
potential for misuse. _Int Arch Med_ 3: 1. Article PubMed PubMed Central Google Scholar * Lawlor DA, Smith GD, Ebrahim S (2003). Association of insulin resistance with depression: cross
sectional findings from the British Women's Heart and Health Study. _BMJ_ 327: 1383–1384. Article PubMed PubMed Central Google Scholar * Lee BH, Kim H, Park SH, Kim YK (2006).
Decreased plasma BDNF level in depressive patients. _J Affect Disord_ 101: 239–244. Article CAS PubMed Google Scholar * Lee JH, Wang LC, Yu HH, Lin YT, Yang YH, Chiang BL (2010). Type I
IL-1 receptor (IL-1RI) as potential new therapeutic target for bronchial asthma. _Mediators Inflamm_ 2010: 567351. Article PubMed PubMed Central CAS Google Scholar * Lee JS, Jang DJ,
Lee N, Ko HG, Kim H, Kim YS _et al_ (2009). Induction of neuronal vascular endothelial growth factor expression by cAMP in the dentate gyrus of the hippocampus is required for
antidepressant-like behaviors. _J Neurosci_ 29: 8493–8505. Article CAS PubMed PubMed Central Google Scholar * Le-Niculescu H, Kurian SM, Yehyawi N, Dike C, Patel SD, Edenberg HJ _et al_
(2009). Identifying blood biomarkers for mood disorders using convergent functional genomics. _Mol Psychiatry_ 14: 156–174. Article CAS PubMed Google Scholar * Leuchter AF, Cook IA,
Hamilton SP, Narr KL, Toga A, Hunter AM _et al_ (2010). Biomarkers to predict antidepressant response. _Curr Psychiatry Rep_ 12: 553–562. Article PubMed PubMed Central Google Scholar *
Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N (1989). Vascular endothelial growth factor is a secreted angiogenic mitogen. _Science_ 246: 1306–1309. Article CAS PubMed Google
Scholar * Levine J, Barak Y, Chengappa KN, Rapoport A, Rebey M, Barak V (1999). Cerebrospinal cytokine levels in patients with acute depression. _Neuropsychobiology_ 40: 171–176. Article
CAS PubMed Google Scholar * Lin E, Chen PS (2008). Pharmacogenomics with antidepressants in the STAR*D study. _Pharmacogenomics_ 9: 935–946. Article CAS PubMed Google Scholar *
Linthorst AC, Flachskamm C, Holsboer F, Reul JM (1994). Local administration of recombinant human interleukin-1 beta in the rat hippocampus increases serotonergic neurotransmission,
hypothalamic–pituitary–adrenocortical axis activity, and body temperature. _Endocrinology_ 135: 520–532. Article CAS PubMed Google Scholar * Liu J, Garza JC, Bronner J, Kim CS, Zhang W,
Lu XY (2010). Acute administration of leptin produces anxiolytic-like effects: a comparison with fluoxetine. _Psychopharmacology_ 207: 535–545. Article CAS PubMed Google Scholar * Lohoff
FW (2010). Overview of the genetics of major depressive disorder. _Curr Psychiatry Rep_ 12: 539–546. Article PubMed PubMed Central Google Scholar * Lommatzsch M, Braun A, Mannsfeldt A,
Botchkarev VA, Botchkareva NV, Paus R _et al_ (1999). Abundant production of brain-derived neurotrophic factor by adult visceral epithelia. Implications for paracrine and target-derived
Neurotrophic functions. _Am J Pathol_ 155: 1183–1193. Article CAS PubMed PubMed Central Google Scholar * Lommatzsch M, Schloetcke K, Klotz J, Schuhbaeck K, Zingler D, Zingler C _et al_
(2005a). Brain-derived neurotrophic factor in platelets and airflow limitation in asthma. _Am J Respir Crit Care Med_ 171: 115–120. Article PubMed Google Scholar * Lommatzsch M, Zingler
D, Schuhbaeck K, Schloetcke K, Zingler C, Schuff-Werner P _et al_ (2005b). The impact of age, weight and gender on BDNF levels in human platelets and plasma. _Neurobiol Aging_ 26: 115–123.
Article CAS PubMed Google Scholar * Lu B, Pang PT, Woo NH (2005). The yin and yang of neurotrophin action. _Nat Rev Neurosci_ 6: 603–614. Article CAS PubMed Google Scholar * Lu XY
(2007). The leptin hypothesis of depression: a potential link between mood disorders and obesity? _Curr Opin Pharmacol_ 7: 648–652. Article CAS PubMed PubMed Central Google Scholar * Lu
XY, Kim CS, Frazer A, Zhang W (2006). Leptin: a potential novel antidepressant. _Proc Natl Acad Sci USA_ 103: 1593–1598. Article CAS PubMed PubMed Central Google Scholar * Lutter M,
Nestler EJ (2009). Homeostatic and hedonic signals interact in the regulation of food intake. _J Nutr_ 139: 629–632. Article CAS PubMed PubMed Central Google Scholar * Lutter M, Sakata
I, Osborne-Lawrence S, Rovinsky SA, Anderson JG, Jung S _et al_ (2008). The orexigenic hormone ghrelin defends against depressive symptoms of chronic stress. _Nat Neurosci_ 11: 752–753.
Article CAS PubMed PubMed Central Google Scholar * Maes M, Bosmans E, De Jongh R, Kenis G, Vandoolaeghe E, Neels H (1997). Increased serum IL-6 and IL-1 receptor antagonist
concentrations in major depression and treatment resistant depression. _Cytokine_ 9: 853–858. Article CAS PubMed Google Scholar * Maier SF, Watkins LR (1995). Intracerebroventricular
interleukin-1 receptor antagonist blocks the enhancement of fear conditioning and interference with escape produced by inescapable shock. _Brain Res_ 695: 279–282. Article CAS PubMed
Google Scholar * Malberg JE, Platt B, Rizzo SJ, Ring RH, Lucki I, Schechter LE _et al_ (2007). Increasing the levels of insulin-like growth factor-I by an IGF binding protein inhibitor
produces anxiolytic and antidepressant-like effects. _Neuropsychopharmacology_ 32: 2360–2368. Article CAS PubMed Google Scholar * Malinowski DP (2007a). Multiple biomarkers in molecular
oncology. I. Molecular diagnostics applications in cervical cancer detection. _Expert Rev Mol Diagn_ 7: 117–131. Article CAS PubMed Google Scholar * Malinowski DP (2007b). Multiple
biomarkers in molecular oncology. II. Molecular diagnostics applications in breast cancer management. _Expert Rev Mol Diagn_ 7: 269–280. Article CAS PubMed Google Scholar * Mallei A, Shi
B, Mocchetti I (2002). Antidepressant treatments induce the expression of basic fibroblast growth factor in cortical and hippocampal neurons. _Mol Pharmacol_ 61: 1017–1024. Article CAS
PubMed Google Scholar * Marrero JA, Kudo M, Bronowicki JP (2010). The challenge of prognosis and staging for hepatocellular carcinoma. _Oncologist_ 15 (Suppl 4): 23–33. Article CAS
PubMed Google Scholar * McIntyre RS, Park KY, Law CW, Sultan F, Adams A, Lourenco MT _et al_ (2010). The association between conventional antidepressants and the metabolic syndrome: a
review of the evidence and clinical implications. _CNS Drugs_ 24: 741–753. Article PubMed Google Scholar * Miller AH, Maletic V, Raison CL (2009). Inflammation and its discontents: the
role of cytokines in the pathophysiology of major depression. _Biol Psychiatry_ 65: 732–741. Article CAS PubMed PubMed Central Google Scholar * Molteni R, Fumagalli F, Magnaghi V,
Roceri M, Gennarelli M, Racagni G _et al_ (2001). Modulation of fibroblast growth factor-2 by stress and corticosteroids: from developmental events to adult brain plasticity. _Brain Res
Brain Res Rev_ 37: 249–258. Article CAS PubMed Google Scholar * Monje ML, Toda H, Palmer TD (2003). Inflammatory blockade restores adult hippocampal neurogenesis. _Science_ 302:
1760–1765. Article CAS PubMed Google Scholar * Monteggia LM, Barrot M, Powell CM, Berton O, Galanis V, Gemelli T _et al_ (2004). Essential role of brain-derived neurotrophic factor in
adult hippocampal function. _Proc Natl Acad Sci USA_ 101: 10827–10832. Article CAS PubMed PubMed Central Google Scholar * Mossner R, Mikova O, Koutsilieri E, Saoud M, Ehlis AC, Muller N
_et al_ (2007). Consensus paper of the WFSBP Task Force on Biological Markers: biological markers in depression. _World J Biol Psychiatry_ 8: 141–174. Article PubMed Google Scholar *
Muller N, Schwarz MJ, Dehning S, Douhe A, Cerovecki A, Goldstein-Muller B _et al_ (2006). The cyclooxygenase-2 inhibitor celecoxib has therapeutic effects in major depression: results of a
double-blind, randomized, placebo controlled, add-on pilot study to reboxetine. _Mol Psychiatry_ 11: 680–684. Article CAS PubMed Google Scholar * Murray CA, Lynch MA (1998). Evidence
that increased hippocampal expression of the cytokine interleukin-1 beta is a common trigger for age- and stress-induced impairments in long-term potentiation. _J Neurosci_ 18: 2974–2981.
Article CAS PubMed PubMed Central Google Scholar * Nakazato M, Hashimoto K, Shimizu E, Kumakiri C, Koizumi H, Okamura N _et al_ (2003). Decreased levels of serum brain-derived
neurotrophic factor in female patients with eating disorders. _Biol Psychiatry_ 54: 485–490. Article CAS PubMed Google Scholar * Nassenstein C, Braun A, Erpenbeck VJ, Lommatzsch M,
Schmidt S, Krug N _et al_ (2003). The neurotrophins nerve growth factor, brain-derived neurotrophic factor, neurotrophin-3, and neurotrophin-4 are survival and activation factors for
eosinophils in patients with allergic bronchial asthma. _J Exp Med_ 198: 455–467. Article CAS PubMed PubMed Central Google Scholar * Nemeroff CB, Musselman DL, Evans DL (1998).
Depression and cardiac disease. _Depression Anxiety_ 8 (Suppl 1): 71–79. Article PubMed Google Scholar * Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM (2002).
Neurobiology of depression. _Neuron_ 34: 13–25. Article CAS PubMed Google Scholar * Newton SS, Collier EF, Hunsberger J, Adams D, Terwilliger R, Selvanayagam E _et al_ (2003). Gene
profile of electroconvulsive seizures: induction of neurotrophic and angiogenic factors. _J Neurosci_ 23: 10841–10851. Article CAS PubMed PubMed Central Google Scholar * Nguyen KT, Deak
T, Owens SM, Kohno T, Fleshner M, Watkins LR _et al_ (1998). Exposure to acute stress induces brain interleukin-1beta protein in the rat. _J Neurosci_ 18: 2239–2246. Article CAS PubMed
PubMed Central Google Scholar * Nibuya M, Morinobu S, Duman RS (1995). Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug
treatments. _J Neurosci_ 15: 7539–7547. Article CAS PubMed PubMed Central Google Scholar * Nibuya M, Takahashi M, Russell DS, Duman RS (1999). Repeated stress increases catalytic TrkB
mRNA in rat hippocampus. _Neurosci Lett_ 267: 81–84. Article CAS PubMed Google Scholar * O’Connor JC, McCusker RH, Strle K, Johnson RW, Dantzer R, Kelley KW (2008). Regulation of IGF-I
function by proinflammatory cytokines: at the interface of immunology and endocrinology. _Cell Immunol_ 252: 91–110. Article PubMed PubMed Central CAS Google Scholar * Okamoto T,
Yoshimura R, Ikenouchi-Sugita A, Hori H, Umene-Nakano W, Inoue Y _et al_ (2008). Efficacy of electroconvulsive therapy is associated with changing blood levels of homovanillic acid and
brain-derived neurotrophic factor (BDNF) in refractory depressed patients: a pilot study. _Prog Neuropsychopharmacol Biol Psychiatry_ 32: 1185–1190. Article CAS PubMed Google Scholar *
Pace TW, Hu F, Miller AH (2007). Cytokine-effects on glucocorticoid receptor function: relevance to glucocorticoid resistance and the pathophysiology and treatment of major depression.
_Brain Behav Immun_ 21: 9–19. Article CAS PubMed Google Scholar * Pan A, Ye X, Franco OH, Li H, Yu Z, Zou S _et al_ (2008). Insulin resistance and depressive symptoms in middle-aged and
elderly Chinese: findings from the Nutrition and Health of Aging Population in China Study. _J Affect Disord_ 109: 75–82. Article CAS PubMed Google Scholar * Pan W, Banks WA, Fasold MB,
Bluth J, Kastin AJ (1998). Transport of brain-derived neurotrophic factor across the blood–brain barrier. _Neuropharmacology_ 37: 1553–1561. Article CAS PubMed Google Scholar * Pan W,
Kastin AJ (2000). Interactions of IGF-1 with the blood-brain barrier _in vivo_ and _in situ_. _Neuroendocrinology_ 72: 171–178. Article CAS PubMed Google Scholar * Papakostas GI, Fava M
(2008). Predictors, moderators, and mediators (correlates) of treatment outcome in major depressive disorder. _Dialogues Clin Neurosci_ 10: 439–451. PubMed PubMed Central Google Scholar *
Papakostas GI, Iosifescu DV, Renshaw PF, Lyoo IK, Lee HK, Alpert JE _et al_ (2005). Brain MRI white matter hyperintensities and one-carbon cycle metabolism in non-geriatric outpatients with
major depressive disorder (Part II). _Psychiatry Res_ 140: 301–307. Article CAS PubMed Google Scholar * Pardridge WM (2002). Neurotrophins, neuroprotection and the blood–brain barrier.
_Curr Opin Investig Drugs_ 3: 1753–1757. CAS PubMed Google Scholar * Pariante CM (2009). Risk factors for development of depression and psychosis. Glucocorticoid receptors and pituitary
implications for treatment with antidepressant and glucocorticoids. _Ann NY Acad Sci_ 1179: 144–152. Article CAS PubMed Google Scholar * Pariante CM, Miller AH (2001). Glucocorticoid
receptors in major depression: relevance to pathophysiology and treatment. _Biol Psychiatry_ 49: 391–404. Article CAS PubMed Google Scholar * Pearson TA, Mensah GA, Alexander RW,
Anderson JL, Cannon III RO, Criqui M _et al_ (2003). Markers of inflammation and cardiovascular disease: application to clinical and public health practice: a statement for healthcare
professionals from the Centers for Disease Control and Prevention and the American Heart Association. _Circulation_ 107: 499–511. Article PubMed Google Scholar * Perlis RH (2011). Betting
on biomarkers. _Am J Psychiatry_ 168: 234–236. Article PubMed PubMed Central Google Scholar * Pittenger C, Duman RS (2007). Stress, depression, and neuroplasticity: a convergence of
mechanisms. _Neuropsychopharmacology_ 33: 88–109. Article PubMed CAS Google Scholar * Poduslo JF, Curran GL (1996). Permeability at the blood–brain and blood–nerve barriers of the
neurotrophic factors: NGF, CNTF, NT-3, BDNF. _Brain Res Mol Brain Res_ 36: 280–286. Article CAS PubMed Google Scholar * Pulford BE, Ishii DN (2001). Uptake of circulating insulin-like
growth factors (IGFs) into cerebrospinal fluid appears to be independent of the IGF receptors as well as IGF-binding proteins. _Endocrinology_ 142: 213–220. Article CAS PubMed Google
Scholar * Qiuhua S, Bergquist-Beringer S, Sousa VD (2011). Major depressive disorder and insulin resistance in nondiabetic young adults in the United States: the National Health and
Nutrition Examination Survey, 1999–2002. _Biol Res Nurs_ 13: 175–181. Article Google Scholar * Quilty LC, Meusel LA, Bagby RM (2008). Neuroticism as a mediator of treatment response to
SSRIs in major depressive disorder. _J Affect Disord_ 111: 67–73. Article CAS PubMed Google Scholar * Raison CL, Capuron L, Miller AH (2006). Cytokines sing the blues: inflammation and
the pathogenesis of depression. _Trends Immunol_ 27: 24–31. Article CAS PubMed Google Scholar * Raison CL, Miller AH (2003). When not enough is too much: the role of insufficient
glucocorticoid signaling in the pathophysiology of stress-related disorders. _Am J Psychiatry_ 160: 1554–1565. Article PubMed Google Scholar * Rasmusson AM, Shi L, Duman R (2002).
Downregulation of BDNF mRNA in the hippocampal dentate gyrus after re-exposure to cues previously associated with footshock. _Neuropsychopharmacology_ 27: 133–142. Article CAS PubMed
Google Scholar * Reinhardt RR, Bondy CA (1994). Insulin-like growth factors cross the blood–brain barrier. _Endocrinology_ 135: 1753–1761. Article CAS PubMed Google Scholar * Rhen T,
Cidlowski JA (2005). Antiinflammatory action of glucocorticoids—new mechanisms for old drugs. _N Engl J Med_ 353: 1711–1723. Article CAS PubMed Google Scholar * Rivier C (1993). Effect
of peripheral and central cytokines on the hypothalamic–pituitary–adrenal axis of the rat. _Ann NY Acad Sci_ 697: 97–105. Article CAS PubMed Google Scholar * Rosa AR, Frey BN, Andreazza
AC, Cereser KM, Cunha AB, Quevedo J _et al_ (2006). Increased serum glial cell line-derived neurotrophic factor immunocontent during manic and depressive episodes in individuals with bipolar
disorder. _Neurosci Lett_ 407: 146–150. Article CAS PubMed Google Scholar * Russo-Neustadt A, Beard RC, Cotman CW (1999). Exercise, antidepressant medications, and enhanced brain
derived neurotrophic factor expression. _Neuropsychopharmacology_ 21: 679–682. Article CAS PubMed Google Scholar * Russo-Neustadt A, Ha T, Ramirez R, Kesslak JP (2001). Physical
activity–antidepressant treatment combination: impact on brain-derived neurotrophic factor and behavior in an animal model. _Behav Brain Res_ 120: 87–95. Article CAS PubMed Google Scholar
* Saarelainen T, Hendolin P, Lucas G, Koponen E, Sairanen M, MacDonald E _et al_ (2003). Activation of the TrkB neurotrophin receptor is induced by antidepressant drugs and is required for
antidepressant-induced behavioral effects. _J Neurosci_ 23: 349–357. Article CAS PubMed PubMed Central Google Scholar * Sairanen M, Lucas G, Ernfors P, Castren M, Castren E (2005).
Brain-derived neurotrophic factor and antidepressant drugs have different but coordinated effects on neuronal turnover, proliferation, and survival in the adult dentate gyrus. _J Neurosci_
25: 1089–1094. Article CAS PubMed PubMed Central Google Scholar * Saito S, Watanabe K, Hashimoto E, Saito T (2009). Low serum BDNF and food intake regulation: a possible new explanation
of the pathophysiology of eating disorders. _Prog Neuropsychopharmacol Biol Psychiatry_ 33: 312–316. Article CAS PubMed Google Scholar * Sales AJ, Biojone C, Terceti MS, Guimaraes FS,
Gomes MV, Joca SR (2011). Antidepressant-like effect induced by systemic and intra-hippocampal administration of DNA methylation inhibitors. _Br J Pharmacol_. (e-pub ahead of print) *
Sapolsky R, Rivier C, Yamamoto G, Plotsky P, Vale W (1987). Interleukin-1 stimulates the secretion of hypothalamic corticotropin-releasing factor. _Science_ 238: 522–524. Article CAS
PubMed Google Scholar * Scarisbrick IA, Jones EG, Isackson PJ (1993). Coexpression of mRNAs for NGF, BDNF, and NT-3 in the cardiovascular system of the pre- and postnatal rat. _J Neurosci_
13: 875–893. Article CAS PubMed PubMed Central Google Scholar * Schinder AF, Poo M (2000). The neurotrophin hypothesis for synaptic plasticity. _Trends Neurosci_ 23: 639–645. Article
CAS PubMed Google Scholar * Schmidt HD, Banasr M, Duman RS (2008). Future antidepressant targets: neurotrophic factors and related signaling cascades. _Drug Discov Today Ther Strateg_ 5:
151–156. Article PubMed PubMed Central Google Scholar * Schmidt HD, Duman RS (2007). The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and
animal models of depressive-like behavior. _Behav Pharmacol_ 18: 391–418. Article CAS PubMed Google Scholar * Schmidt HD, Duman RS (2010). Peripheral BDNF produces antidepressant-like
effects in cellular and behavioral models. _Neuropsychopharmacology_ 35: 2378–2391. Article CAS PubMed PubMed Central Google Scholar * Schulte-Herbruggen O, Nassenstein C, Lommatzsch M,
Quarcoo D, Renz H, Braun A (2005). Tumor necrosis factor-alpha and interleukin-6 regulate secretion of brain-derived neurotrophic factor in human monocytes. _J Neuroimmunol_ 160: 204–209.
Article PubMed CAS Google Scholar * Sen S, Duman R, Sanacora G (2008). Serum brain-derived neurotrophic factor, depression, and antidepressant medications: meta-analyses and
implications. _Biol Psychiatry_ 64: 527–532. Article CAS PubMed PubMed Central Google Scholar * Shelton RC, Miller AH (2010). Eating ourselves to death (and despair): the contribution
of adiposity and inflammation to depression. _Prog Neurobiol_ 91: 275–299. Article CAS PubMed PubMed Central Google Scholar * Shimizu E, Hashimoto K, Okamura N, Koike K, Komatsu N,
Kumakiri C _et al_ (2003). Alterations of serum levels of brain-derived neurotrophic factor (BDNF) in depressed patients with or without antidepressants. _Biol Psychiatry_ 54: 70–75. Article
CAS PubMed Google Scholar * Shirayama Y, Chen AC, Nakagawa S, Russell DS, Duman RS (2002). Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of
depression. _J Neurosci_ 22: 3251–3261. Article CAS PubMed PubMed Central Google Scholar * Simen BB, Duman CH, Simen AA, Duman RS (2006). TNFalpha signaling in depression and anxiety:
behavioral consequences of individual receptor targeting. _Biol Psychiatry_ 59: 775–785. Article CAS PubMed Google Scholar * Siuciak JA, Lewis DR, Wiegand SJ, Lindsay RM (1997).
Antidepressant-like effect of brain-derived neurotrophic factor (BDNF). _Pharmacol Biochem Behav_ 56: 131–137. Article CAS PubMed Google Scholar * Skilton MR, Moulin P, Terra JL, Bonnet
F (2007). Associations between anxiety, depression, and the metabolic syndrome. _Biol Psychiatry_ 62: 1251–1257. Article CAS PubMed Google Scholar * Sluzewska A, Rybakowski J, Bosmans E,
Sobieska M, Berghmans R, Maes M _et al_ (1996). Indicators of immune activation in major depression. _Psychiatry Res_ 64: 161–167. Article CAS PubMed Google Scholar * Smith MA, Makino
S, Altemus M, Michelson D, Hong SK, Kvetnansky R _et al_ (1995a). Stress and antidepressants differentially regulate neurotrophin 3 mRNA expression in the locus coeruleus. _Proc Natl Acad
Sci USA_ 92: 8788–8792. Article CAS PubMed PubMed Central Google Scholar * Smith MA, Makino S, Kvetnansky R, Post RM (1995b). Stress and glucocorticoids affect the expression of
brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. _J Neurosci_ 15 (3 Pt 1): 1768–1777. Article CAS PubMed PubMed Central Google Scholar * Song C, Halbreich
U, Han C, Leonard BE, Luo H (2009). Imbalance between pro- and anti-inflammatory cytokines, and between Th1 and Th2 cytokines in depressed patients: the effect of electroacupuncture or
fluoxetine treatment. _Pharmacopsychiatry_ 42: 182–188. Article CAS PubMed Google Scholar * Steptoe A, Hamer M, Chida Y (2007). The effects of acute psychological stress on circulating
inflammatory factors in humans: a review and meta-analysis. _Brain Behav Immun_ 21: 901–912. Article CAS PubMed Google Scholar * Stewart CE, Rotwein P (1996). Growth, differentiation,
and survival: multiple physiological functions for insulin-like growth factors. _Physiol Rev_ 76: 1005–1026. Article CAS PubMed Google Scholar * Storkebaum E, Lambrechts D, Carmeliet P
(2004). VEGF: once regarded as a specific angiogenic factor, now implicated in neuroprotection. _Bioessays_ 26: 943–954. Article CAS PubMed Google Scholar * Sutcigil L, Oktenli C,
Musabak U, Bozkurt A, Cansever A, Uzun O _et al_ (2007). Pro- and anti-inflammatory cytokine balance in major depression: effect of sertraline therapy. _Clin Dev Immunol_ 2007: 76396.
Article PubMed Google Scholar * Takebayashi M, Hisaoka K, Nishida A, Tsuchioka M, Miyoshi I, Kozuru T _et al_ (2006). Decreased levels of whole blood glial cell line-derived neurotrophic
factor (GDNF) in remitted patients with mood disorders. _Int J Neuropsychopharmacol_ 9: 607–612. Article CAS PubMed Google Scholar * Taler M, Gil-Ad I, Lomnitski L, Korov I, Baharav E,
Bar M _et al_ (2007). Immunomodulatory effect of selective serotonin reuptake inhibitors (SSRIs) on human T lymphocyte function and gene expression. _Eur Neuropsychopharmacol_ 17: 774–780.
Article CAS PubMed Google Scholar * Taliaz D, Loya A, Gersner R, Haramati S, Chen A, Zangen A (2011). Resilience to chronic stress is mediated by hippocampal brain-derived neurotrophic
factor. _J Neurosci_ 31: 4475–4483. Article CAS PubMed PubMed Central Google Scholar * Thomas AJ, Davis S, Morris C, Jackson E, Harrison R, O’Brien JT (2005). Increase in
interleukin-1beta in late-life depression. _Am J Psychiatry_ 162: 175–177. Article PubMed Google Scholar * Timmusk T, Palm K, Metsis M, Reintam T, Paalme V, Saarma M _et al_ (1993).
Multiple promoters direct tissue-specific expression of the rat BDNF gene. _Neuron_ 10: 475–489. Article CAS PubMed Google Scholar * Timonen M, Laakso M, Jokelainen J, Rajala U,
Meyer-Rochow VB, Keinanen-Kiukaanniemi S (2005). Insulin resistance and depression: cross sectional study. _BMJ_ 330: 17–18. Article PubMed PubMed Central Google Scholar * Timonen M,
Rajala U, Jokelainen J, Keinanen-Kiukaanniemi S, Meyer-Rochow VB, Rasanen P (2006). Depressive symptoms and insulin resistance in young adult males: results from the Northern Finland 1966
birth cohort. _Mol Psychiatry_ 11: 929–933. Article CAS PubMed Google Scholar * Toyooka K, Asama K, Watanabe Y, Muratake T, Takahashi M, Someya T _et al_ (2002). Decreased levels of
brain-derived neurotrophic factor in serum of chronic schizophrenic patients. _Psychiatry Res_ 110: 249–257. Article CAS PubMed Google Scholar * Trejo JL, Carro E, Torres-Aleman I
(2001). Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. _J Neurosci_ 21: 1628–1634. Article CAS PubMed
PubMed Central Google Scholar * Trejo JL, Llorens-Martin MV, Torres-Aleman I (2008). The effects of exercise on spatial learning and anxiety-like behavior are mediated by an
IGF-I-dependent mechanism related to hippocampal neurogenesis. _Mol Cell Neurosci_ 37: 402–411. Article CAS PubMed Google Scholar * Trejo JL, Piriz J, Llorens-Martin MV, Fernandez AM,
Bolos M, LeRoith D _et al_ (2007). Central actions of liver-derived insulin-like growth factor I underlying its pro-cognitive effects. _Mol Psychiatry_ 12: 1118–1128. Article CAS PubMed
Google Scholar * Tsankova NM, Berton O, Renthal W, Kumar A, Neve RL, Nestler EJ (2006). Sustained hippocampal chromatin regulation in a mouse model of depression and antidepressant action.
_Nat Neurosci_ 9: 519–525. Article CAS PubMed Google Scholar * Tuglu C, Kara SH, Caliyurt O, Vardar E, Abay E (2003). Increased serum tumor necrosis factor-alpha levels and treatment
response in major depressive disorder. _Psychopharmacology (Berl)_ 170: 429–433. Article CAS Google Scholar * Tyring S, Gottlieb A, Papp K, Gordon K, Leonardi C, Wang A _et al_ (2006).
Etanercept and clinical outcomes, fatigue, and depression in psoriasis: double-blind placebo-controlled randomised phase III trial. _Lancet_ 367: 29–35. Article CAS PubMed Google Scholar
* Vauthey JN, Dixon E, Abdalla EK, Helton WS, Pawlik TM, Taouli B _et al_ (2010). Pretreatment assessment of hepatocellular carcinoma: expert consensus statement. _HPB (Oxford)_ 12:
289–299. Article Google Scholar * Ventriglia M, Zanardini R, Pedrini L, Placentino A, Nielsen MG, Gennarelli M _et al_ (2009). VEGF serum levels in depressed patients during SSRI
antidepressant treatment. _Prog Neuropsychopharmacol Biol Psychiatry_ 33: 146–149. Article CAS PubMed Google Scholar * von der Thusen JH, Kuiper J, van Berkel TJ, Biessen EA (2003).
Interleukins in atherosclerosis: molecular pathways and therapeutic potential. _Pharmacol Rev_ 55: 133–166. Article CAS PubMed Google Scholar * Wager TD, Davidson ML, Hughes BL,
Lindquist MA, Ochsner KN (2008). Prefrontal-subcortical pathways mediating successful emotion regulation. _Neuron_ 59: 1037–1050. Article CAS PubMed PubMed Central Google Scholar *
Wallace DL, Han MH, Graham DL, Green TA, Vialou V, Iniguez SD _et al_ (2009). CREB regulation of nucleus accumbens excitability mediates social isolation-induced behavioral deficits. _Nat
Neurosci_ 12: 200–209. Article CAS PubMed PubMed Central Google Scholar * Wang PS, Simon G, Kessler RC (2003). The economic burden of depression and the cost-effectiveness of treatment.
_Int J Methods Psychiatr Res_ 12: 22–33. Article CAS PubMed Google Scholar * Warner-Schmidt JL, Duman RS (2007). VEGF is an essential mediator of the neurogenic and behavioral actions
of antidepressants. _Proc Natl Acad Sci USA_ 104: 4647–4652. Article CAS PubMed PubMed Central Google Scholar * Weaver IC, Cervoni N, Champagne FA, D’Alessio AC, Sharma S, Seckl JR _et
al_ (2004). Epigenetic programming by maternal behavior. _Nat Neurosci_ 7: 847–854. Article CAS PubMed Google Scholar * Werther GA, Abate M, Hogg A, Cheesman H, Oldfield B, Hards D _et
al_ (1990). Localization of insulin-like growth factor-I mRNA in rat brain by _in situ_ hybridization—relationship to IGF-I receptors. _Mol Endocrinol_ 4: 773–778. Article CAS PubMed
Google Scholar * Wong ML, Kling MA, Munson PJ, Listwak S, Licinio J, Prolo P _et al_ (2000). Pronounced and sustained central hypernoradrenergic function in major depression with
melancholic features: relation to hypercortisolism and corticotropin-releasing hormone. _Proc Natl Acad Sci USA_ 97: 325–330. Article CAS PubMed PubMed Central Google Scholar * Yamada
N, Katsuura G, Ochi Y, Ebihara K, Kusakabe T, Hosoda K _et al_ (2011). Impaired CNS leptin action is implicated in depression associated with obesity. _Endocrinology_ 152: 2634–2643. Article
CAS PubMed Google Scholar * Yamamoto M, Sobue G, Yamamoto K, Terao S, Mitsuma T (1996). Expression of mRNAs for neurotrophic factors (NGF, BDNF, NT-3, and GDNF) and their receptors
(p75NGFR, trkA, trkB, and trkC) in the adult human peripheral nervous system and nonneural tissues. _Neurochem Res_ 21: 929–938. Article CAS PubMed Google Scholar * Yirmiya R, Pollak Y,
Morag M, Reichenberg A, Barak O, Avitsur R _et al_ (2000). Illness, cytokines, and depression. _Ann NY Acad Sci_ 917: 478–487. Article CAS PubMed Google Scholar * Yoshimura R, Mitoma M,
Sugita A, Hori H, Okamoto T, Umene W _et al_ (2007). Effects of paroxetine or milnacipran on serum brain-derived neurotrophic factor in depressed patients. _Prog Neuropsychopharmacol Biol
Psychiatry_ 31: 1034–1037. Article CAS PubMed Google Scholar * Yurkovetsky Z, Skates S, Lomakin A, Nolen B, Pulsipher T, Modugno F _et al_ (2010). Development of a multimarker assay for
early detection of ovarian cancer. _J Clin Oncol_ 28: 2159–2166. Article PubMed PubMed Central Google Scholar * Zanardini R, Gazzoli A, Ventriglia M, Perez J, Bignotti S, Rossini PM _et
al_ (2006). Effect of repetitive transcranial magnetic stimulation on serum brain derived neurotrophic factor in drug resistant depressed patients. _J Affect Disord_ 91: 83–86. Article CAS
PubMed Google Scholar * Zhu CS, Pinsky PF, Cramer DW, Ransohoff DF, Hartge P, Pfeiffer RM _et al_ (2011). A framework for evaluating biomarkers for early detection: validation of
biomarker panels for ovarian cancer. _Cancer Prevent Res_ 4: 375–383. Article Google Scholar Download references ACKNOWLEDGEMENTS We thank Dr Andrew H Miller for critical reading of the
manuscript. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Psychiatry, Center for Neurobiology and Behavior, University of Pennsylvania School of Medicine, Philadelphia, PA, USA
Heath D Schmidt * Department of Psychiatry, Vanderbilt University School of Medicine, Nashville, TN, USA Richard C Shelton * Department of Psychiatry, Laboratory of Molecular Psychiatry,
Center for Genes and Behavior, Yale University School of Medicine, Connecticut Mental Health Center, New Haven, CT, USA Ronald S Duman * Department of Pharmacology, Laboratory of Molecular
Psychiatry, Center for Genes and Behavior, Yale University School of Medicine, Connecticut Mental Health Center, New Haven, CT, USA Ronald S Duman Authors * Heath D Schmidt View author
publications You can also search for this author inPubMed Google Scholar * Richard C Shelton View author publications You can also search for this author inPubMed Google Scholar * Ronald S
Duman View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to Heath D Schmidt. ETHICS DECLARATIONS COMPETING INTERESTS Drs
Heath D Schmidt and Ronald S Duman report no biomedical financial interests or potential conflicts of interest. Richard C Shelton, MD, has served as a consultant for Eli Lilly &
Company, Evotec AG, Forest Pharmaceuticals, Janssen Pharmaceutica, Medtronic Inc., Otsuka Pharmaceuticals, Pamlab Inc., Pfizer Inc., Repligen Inc., and Ridge Diagnostics. He has received
research support from Eli Lilly & Company, Euthymics Bioscience, Forest Pharmaceuticals, Janssen Pharmaceutica, Novartis Pharmaceuticals, Otsuka Pharmaceuticals, Pamlab, Pfizer Inc.,
Repligen Inc., Ridge Diagnostics, and St Jude Medical. POWERPOINT SLIDES POWERPOINT SLIDE FOR FIG. 1 POWERPOINT SLIDE FOR FIG. 2 RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS
ARTICLE CITE THIS ARTICLE Schmidt, H., Shelton, R. & Duman, R. Functional Biomarkers of Depression: Diagnosis, Treatment, and Pathophysiology. _Neuropsychopharmacol_ 36, 2375–2394
(2011). https://doi.org/10.1038/npp.2011.151 Download citation * Received: 18 April 2011 * Revised: 13 June 2011 * Accepted: 05 July 2011 * Published: 03 August 2011 * Issue Date: November
2011 * DOI: https://doi.org/10.1038/npp.2011.151 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is
not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * antidepressant * BDNF * IGF-1 * IL-1_β_ * serum *
mood disorders