Play all audios:
ABSTRACT Ferroptosis is a recently discovered essential type of cell death that is mainly characterized by iron overload and lipid peroxidation. Emerging evidence suggests that ferroptosis
is a double-edged sword in human cancer. However, the precise underlying molecular mechanisms and their differential roles in tumorigenesis are unclear. Therefore, in this review, we
summarize and briefly present the key pathways of ferroptosis, paying special attention to the regulation of ferroptosis as well as its dual role as an oncogenic and as a tumor suppressor
event in various human cancers. Moreover, multiple pharmacological ferroptosis activators are summarized, and the prospect of targeting ferroptosis in cancer therapy is further elucidated.
SIMILAR CONTENT BEING VIEWED BY OTHERS FERROPTOSIS: A DOUBLE-EDGED SWORD Article Open access 30 May 2024 FERROPTOSIS IN CANCER: FROM MOLECULAR MECHANISMS TO THERAPEUTIC STRATEGIES Article
Open access 08 March 2024 THE DIVERSIFIED ROLE OF MITOCHONDRIA IN FERROPTOSIS IN CANCER Article Open access 14 August 2023 FACTS * 1. Ferroptosis often dysregulated in human cancer. * 2.
Ferroptosis plays a dual role in cancer. * 3. Ferroptosis provides a promising strategy for cancer therapy. OPEN QUESTIONS * 1. Why ferroptosis is a double-edged sword in cancer and what is
its mechanism? * 2. What is the mechanism of ncRNAs regulating ferroptosis and whether they can be the targets of ferroptosis? * 3. In clinical treatment, how to combine ferroptosis inducers
with anti-tumor therapy to achieve optimal therapeutic effect? INTRODUCTION In 2020, there were an estimated 19.3 million new cancer cases and nearly 10 million mortalities due to cancer
worldwide [1]. Despite significant advances in treatment, cancer remains the leading cause of death in humans. Furthermore, toxic effects and drug resistance of traditional therapies pose
major challenges. As a result, the search for more effective and tolerable anticancer therapies continues [2]. Ferroptosis was first defined by Brent R. Stockwell in 2012 as an
iron-dependent type of cell death that is driven by lipid reactive oxygen species (ROS) [3,4,5] and is significantly different from other regulated cell death forms such as apoptosis [6, 7],
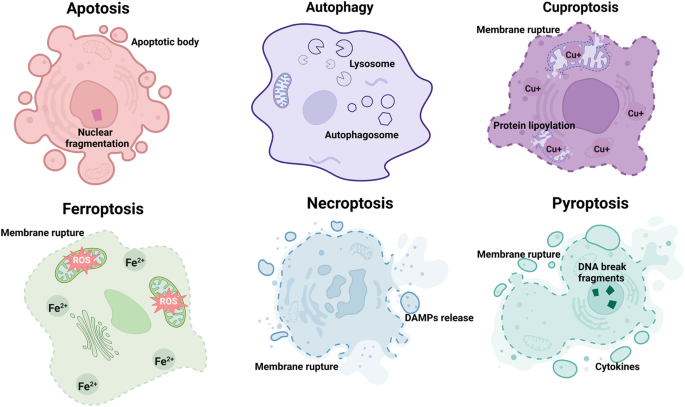
autophagy [8, 9], cuproptosis [10, 11], necroptosis [12, 13], and pyroptosis [13, 14] in terms of cell morphology, biochemistry, and genetics (Fig. 1, Fig. 2 and Table 1). Both the
extrinsic pathway and the intrinsic pathway play a crucial role in this process. On the one hand, the extrinsic pathway is activation by inhibition of the cystine-glutamate antiporter
(system XC—). The absorption of extracellular cystine is hindered by the decreased activity of system XC—, which subsequently affects the synthesis of glutathione (GSH) [15]. This reduction
in the cell’s antioxidant capacity can result in the accumulation of lipid ROS, thus promoting ferroptosis [15, 16]. In contrast, activation of iron transporters such as serum transferrin
and lactotransferrin could trigger the extrinsic pathway. By increasing iron intake and limiting iron loss, these transporters induce ferroptosis via enhancing iron accumulation [15, 17].
The intrinsic pathway is realized by blocking the activation of intracellular antioxidant enzymes. For example, GPX4 can reduce the cytotoxic lipid peroxide (L-OOH) to the corresponding
alcohol (L-OH). Once the activity of GPX4 is inhibited, it will lead to the accumulation of lipid peroxide in the cell membrane [15, 18]. In addition, long-chain fatty acid coenzyme A ligase
4 (ACSL4) encourage the incorporation of polyunsaturated fatty acids (PUFA) into phospholipids to form polyunsaturated fatty acid phospholipids (PUFA-PL). PUFA-PLs are susceptible to
oxidation induced by free radicals mediated by lipoxygenases (ALOXs), which eventually lead to the destruction of the lipid bilayer and affect the membrane function, thus triggering
ferroptosis [19, 20]. Both extrinsic and intrinsic pathways can be directly activated by specific small molecules, such as RAS-selective lethal (RSL3) and erastin. Therefore, the selection
of small molecules for extrinsic or intrinsic pathways can be based on the level of development difficulty. However, some small molecules such as ferristatin 1 are not suitable for in vivo
experiments due to its solubility, stability, pharmacokinetics, and drug delivery route, which is a major challenge in clinical transformation. Recently, ferroptosis has been shown to be
involved in different diseases including neurodegeneration and malignant tumors such as liver cancer, breast cancer, and lung cancer [21,22,23,24]. Over the past two decades, ferroptosis has
been extensively studied in a variety of malignancies, and it has been shown to regulate cancer progression through various signaling pathways. Fascinatingly, it is not as only a tumor
suppressor, but it also exerts oncogenic function. Of note, cancer cells that are resistant to traditional therapies or have a high propensity to metastasize are more prone to ferroptosis
[25, 26]. Hence, the regulation of ferroptosis and its target proteins is considered a novel and promising cancer therapy strategy. The multiple signaling pathways involved in ferroptosis
have been well summarized elsewhere [27, 28]. Herein, the purpose of this article is to provide insights into the involvement of ferroptosis, along with its regulators and promoters, in
tumor development and as a potential treatment target. In particular, potential applications of ferroptosis in chemotherapy, radiotherapy, immunotherapy, nanotherapy, sonodynamic therapy
(SDT) and photodynamic therapy (PDT) are discussed, with an emphasis on potential therapeutic agents for clinical applications. FERROPTOSIS IS AN ESSENTIAL TYPE OF CELL DEATH Ferroptosis is
defined as a form of regulated cell death that is characterized by the accumulation of iron and ROS [3,4,5]. Specifically, ROS is highly active oxidation radical, including superoxides,
peroxides and free radicals [29, 30]. Fenton reaction is an important source of ROS, which is produced by the interaction of ferrous iron ion and hydrogen peroxide in cells through Fenton
reaction [30, 31]. Subsequently, ROS reacts with PUFAs to deprive the hydrogen atom between its long chain double bonds, causing lipid peroxidation and ultimately leading to ferroptosis [31,
32]. On the other hand, mitochondria are another essential source of ROS. During oxidative phosphorylation of mitochondria, ROS is produced after the combination of electrons and molecular
oxygen, which increases the oxidative stress response of cells, accompanied by lipid, protein, and DNA damage [31,32,33]. This will lead to accumulation of lipid ROS, atrophy of
mitochondria, increase of cell membrane density, and eventually result in ferroptosis. Interestingly, the signaling mechanisms of ferroptosis and other cell death types of crosstalk with
each other. For example, ROS is essential for autophagy, apoptosis, and ferroptosis; in addition, the core regulator of ferroptosis, GSH, is also involved in the regulation of cuproptosis
[34, 35]. Nonetheless, ferroptosis has been reported to induce specific cellular morphological and biochemical changes and to be regulated by multiple metabolic pathways (Figs. 1, 2 and
Table 1). MORPHOLOGICAL AND BIOCHEMICAL FEATURES OF FERROPTOSIS The morphological features of cells undergoing ferroptosis differ from those of cells undergoing autophagy or apoptosis.
Specifically, the cell membrane is not damaged during apoptosis or autophagy; while in ferroptosis, the cell membrane density is increased and ruptured, leading to the formation of vesicles.
Additionally, the hallmarks of necroptosis or pyroptosis are cytoplasmic swelling and DNA breakage, respectively [13, 36]. However, in ferroptosis, the distinctive characteristics are
manifested as mitochondrial atrophy as well as a reduction or even disappearance of the mitochondrial ridge, but there is no change in the size of the nucleus except for chromatin
condensation [24, 37, 38]. Moreover, the signs of apoptosis, pyroptosis, and cuprotosis are the formation of apoptotic bodies, the release of inflammatory cytokines, and copper accumulation;
while ferroptosis is mainly distinguished by an excess of iron and ROS [3,4,5, 10, 39, 40]. In short, ferroptosis and known forms of regulated cell death have distinct morphological and
biochemical features (Fig. 1 and Table 1). METABOLIC CHANGES IN FERROPTOSIS IRON METABOLISM IMBALANCE Iron metabolism is regulated by multiple steps, including iron absorption, utilization,
recycling, and storage. Disruption of iron metabolism leads to excessive accumulation of intracellular iron, which causes the generation of free radicals and oxidative stress [41, 42].
Specifically, as one of the essential elements in the proliferation and development of tumor cells, iron is also the core element for ferroptosis [43, 44]. In addition to its role in the
production of DNA and ATP, iron is a crucial component of the mitochondrial electron transport chain and a cofactor of metalloproteinases. For example, ferrithioprotein is simultaneously a
vital cofactor of oxidoreductases in the mitochondrial electron transport chain and a cofactor of many vital enzymes in redox reactions. Extracellular ferric ions (Fe3+) combine with
transferrin to form the transferrin–Fe3+ complex, which enters the cell mediated by the membrane protein transferrin receptor 1 and is reduced to Fe2+. With the assistance of the divalent
metal transporter 1 (solute carrier family 11 member 2; SLC11A2) or Zrt- and Irt-like proteins 8 and 14 (SLC39A8 and SLC39A14, respectively), the labile iron pool stores Fe2+ in the cell
[43, 45, 46]. Meanwhile, Fe2+, with the help of iron chaperones such as poly(rC)-binding proteins 1 and 2, pumps iron through membrane ferroportin (FPN) 1 to maintain the balance of
intracellular iron [43, 47]. However, once Fe2+ in cells is overloaded, the Fenton reaction will occur with hydrogen peroxide, resulting in excessive ROS and driving ferroptosis (Fig. 2).
LIPID METABOLISM DISORDER Lipids have crucial roles in energy storage, signal transmission, membrane development, cell membrane formation, energy storage, and signal transduction. Lipid
metabolism regulates cell lipid toxicity, and abnormal lipid metabolism is considered a hallmark of malignancy and an essential factor for ferroptosis [48]. Additionally, the metabolism of
lipids in cells relies heavily on fatty acids. According to their saturation levels, fatty acids are divided into three categories: saturated fatty acids, PUFAs, and monounsaturated fatty
acids (MUFAs). Among them, PUFAs and MUFAs have been demonstrated to contribute to ferroptosis [49, 50]. Due to the weak C–H bond at the diallyl position, PUFAs on the cell membrane are
vulnerable to attack by ROS, which can induce lipid peroxidation [51]. In this process, acyl-CoA synthetase long-chain family member 4 (ACSL4) is required for the production of PUFAs,
thereby positively regulating ferroptosis [51]. In contrast, exogenous MUFAs such as exogenous palmitic acid and oleic acid have been reported to negatively regulate drug-induced ferroptosis
[52, 53]. Exogenous MUFAs can be activated by acyl-CoA synthetase long-chain family member 3 to displace PUFAs at the plasma membrane and to reduce the sensitivity of plasma membrane lipids
to oxidation [53]. Moreover, an increased ratio of MUFAs to PUFAs can be observed on cancer cell membranes, thereby inhibiting lipotoxicity and ferroptosis [54] (Fig. 2). AMINO ACID
METABOLISM DYSFUNCTION Amino acids are essential nutrients for the survival of cells and are involved in deamination, decarboxylation, ammonia metabolism, and oxidative decomposition
capacity metabolic processes. Meanwhile, abnormal amino acid metabolism leads to redox imbalance, energy regulation disorder, and biosynthesis dysfunction, thus providing support for tumor
proliferation [55]. Ferroptosis induced by an abnormal amino acid metabolism is mainly related to GSH. GSH is a tripeptide compound composed of glutamate, cysteine, and glycine
(γ-glutamyl-L-cysteinyl-L-glycine) and is an important antioxidant and free radical scavenger in the body [56, 57]. System XC— and GPX4 are key regulators of GSH biosynthesis and
degradation. System XC— is composed of light (SLC7A11) and heavy chain (SLC3A2) subunits, which play an essential role in maintaining the balance of GSH in cells [58, 59]. System XC—
facilitates the interchange of cystine and glutamate across the plasma membrane, controls GSH production in response to extracellular glutamate levels, and transports glutamate from inside
the cell to outside the cell [60]. Impaired function of system XC— or insufficient intracellular cysteine levels can lead to the decreased synthesis of GSH, triggering ferroptosis. On the
other hand, as an important enzyme for scavenging lipid oxygen free radicals, GPX4 can use GSH as a substrate to reduce membrane lipid hydrogen peroxide to nontoxic lipid alcohols, decrease
oxidative stress damage, and negatively regulate ferroptosis [56, 57] (Fig. 2). REGULATION OF FERROPTOSIS IN CANCER Ferroptosis can be regulated by a variety of mechanisms, especially by the
transcription of genes and post-translation of proteins. NONCODING RNAS (NCRNAS) PROMOTE FERROPTOSIS IN CANCER Numerous studies have demonstrated that ncRNAs can promote ferroptosis in
multiple cancers (Table 2). For example, microRNA (miRNA or miR)-15a and miR-15a-3p have been reported to target GPX4 to promote ferroptosis in prostate cancer and colorectal cancer,
respectively [61, 62]. Furthermore, GPX4 inhibition triggers the sensitivity of ferroptosis by restoring the miR-4715-3p levels, which are suppressed in upper gastrointestinal cancers [63].
A similar mechanism has been observed in non-small cell lung cancer. miR-324-3p causes cisplatin resistance by suppressing GPX4 expression [64], while miR-302a-3p acts as a positive
regulator of ferroptosis by targeting FPN [65]. In addition to inducing ferroptosis via GPX4, a study by Bai et al. has shown that miR-214-3p the GSH axis in hepatoma and thus acts as a
tumor inhibitor [66]. Similar to miRNAs, ferroptosis can be accelerated by some long noncoding RNAs (lncRNAs) in various cancers. Taking liver cancer as an example, it has been observed that
knockdown of the lncRNA plasmacytoma variant translocation 1 significantly increased the levels of ROS and Fe2+, followed by inhibition of cell viability [67]. In renal cancer, silencing of
SLC16A1-AS1 reduced the expression of SLC7A11 and significantly decreased the GSH/glutathione disulfide (GSSG) ratio in cells [68]. Similar results were also confirmed by Wang et al., who
demonstrated that the long intergenic nonprotein-coding RNA 618 activates ferroptosis via upregulating SLC7A11 and downregulating ACSL4 in acute myeloid leukemia [69]. Mechanistically, the
lncRNA ARHGEF26-AS1 acts as a sponge for miR-372-3p, not only promoting ferroptosis but also inhibiting the proliferation and migration of esophageal squamous cell carcinoma cells [70].
Consistently, the cytosolic p53-related lncRNA induces ferroptosis via promoting ROS and intracellular iron accumulation, thereby suppressing lung cancer progression [71]. Additionally,
bioinformatics analysis and preclinical experiments have verified that metallothionein 1D, pseudogene enhances erastin-induced ferroptosis by inhibiting nuclear factor erythroid 2-related
factor 2 (NRF2) in nonsmall cell lung cancer [72]. A number of circular RNAs also have been reported to be involved in the promotion of ferroptosis. Specifically, according to research by
Jiang et al., circ0000190 overexpression accelerated ferroptosis in gastric cancer cells by increasing the levels of malondialdehyde, lipid ROS, and Fe2+ [73]. In addition, in colorectal
cancer, after knockdown of circ0007142, cells sent out signals for ferroptosis and growth inhibition was observed [74]. Moreover, a preclinical study has shown that apoptosis-inducing factor
mitochondria-associated 2 and GPX4 expression as well as the GSH/GSSG ratio were upregulated after knockdown of the circular RNA glial cell line-derived neurotrophic factor family receptor
alpha-1 (circGFRA1), suggesting that circGFRA1 promotes ferroptosis via two independent pathways in breast cancer [75]. Of note, experiments performed in vitro and in vivo have revealed that
the circular RNA LIM domain only 1 enhances ferroptosis by increasing the expression of ACSL4, which in turn slows down the growth and metastasis of cervical cancer cells [76]. Furthermore,
according to RNA-sequencing analysis, the circular RNA IARS (circ-IARS) is overexpressed in hepatocellular carcinoma (HCC), while an in-depth study has found that circ-IARS-silenced cells
show a substantial decrease in Fe2+ and an evident increase in intracellular GSH. Therefore, it can be speculated that circ-IARS may act as a catalyst for ferroptosis in HCC cells [77].
However, the relationship between ferroptosis and ncRNAs is largely unknown and there are facing many challenges. For example, more detailed and evidence are needed to elucidate the
underlying regulatory mechanism between ferroptosis and ncRNAs in the future study. To date, there is still no evidence showing that ncRNAs involved in the occurrence and prognosis of cancer
by directly binding to ferroptosis. Therefore, it is essential to further explore more roles of ferroptosis-related ncRNAs in different cancers. In addition, only a fraction of
ferroptosis-related ncRNAs have been validated in vivo. Further investigation in large-scale human tissue samples is obviously warranted to determine whether these ncRNAs can be used as a
target for clinical. TWO SIDES OF TRANSCRIPTIONAL REGULATORS IN FERROPTOSIS Growing evidence has implicated that transcriptional regulators are double-edged swords in ferroptosis regulation.
For example, p53 as a tumor suppressor, has been reported to play a dual role in ferroptosis. In particular, p53 has been demonstrated to induce ferroptosis by directly inhibiting the
expression of SLC7A11 and enhancing lipid peroxidase. Of note, it can be detected that the activation of p53 reduced the uptake of cystine, limited intracellular GSH production, thereby
activated ferroptosis and suppressed tumor growth [78]. However, p53 also has been proposed to play a role in blocking ferroptosis in human cancer cells. Mechanically, the expression of p53
was increased under the treatment with a small molecule inhibitor, nutlin-3, accompanied by the decrease in GSH consumption and ROS accumulation, ultimately suppressing ferroptosis.
Simultaneously, an increase in cell viability can be observed in HT-1080 fibrosarcoma cells [79, 80]. Furthermore, activating transcription factor (ATF) 4 also functions as a negative or
positive regulator of ferroptosis in various cancers. On the one hand, ATF4 can drive sorafenib resistance in HCC by inhibiting ferroptosis [81]; on the other hand, sevoflurane has been
shown to induce ferroptosis of glioma cells through ATF4 activation [82]. Similarly, ATF3 has been reported to promote ferroptosis and to exert tumor-suppressing effects [83]. Moreover, the
abnormal expression of yes-associated protein (YAP) and transcriptional co-activator with PDZ-binding motif (TAZ), two core transcription factors of Hippo pathway, contributes to the cell
growth and chemotherapy resistance in multiple cancers [84, 85]. Furthermore, a preclinical study has demonstrated that transcriptional regulatory activity of YAP triggered ferroptosis by
targeting transferrin receptor and ACSL4. Briefly, overexpression of YAP lead to an increase of ROS and decrease of cell viability. Moreover, YAP is more susceptible to ferroptosis at high
cell density in colon cancer cells [86]. Consistent with this finding, loss of TAZ reduces susceptibility to ferroptosis in multiple cancer cell lines treated with erastin [87]. Besides,
hypoxia-inducible factor 1 alpha (HIF1A), as a transcriptional factor of the homeostatic response of cells to hypoxia, is considered to inhibit cancer cell death by promoting lipid
accumulation. However, in mice models treated with RSL3, knockout of HIF1A was confirmed to positively regulate ferroptosis by regulating lipid metabolism, thereby effectively inhibiting
tumor growth [88, 89]. On the contrary, there is also transcription regulator that promote tumor development by inhibiting ferroptosis. The most direct proof is that NRF2 has been shown to
upregulate SLC7A11, thereby protecting tumor cells from ferroptosis [90]. Nevertheless, NRF2 was also reported to trigger ferroptosis via increasing the expression of HMOX1 in lung cancer
and renal cell carcinoma (RCC) cells. Specifically, NRF2 can be activated by 4,4'-dimethoxychalcone (DMC) which is extracted from the plant _Angelica keiskei koidzumi_. The activation
of NRF2 directly upregulated HMOX1 expression, subsequently led to iron overload and ferroptosis [91, 92] (Fig. 3). Given a dual role of transcription factors play in ferroptosis, it will be
intriguing to further clarify the specific mechanism of this process. It will also be interesting to screen much more transcription factors targeting ferroptosis. In addition, due to the
complex regulatory network of ferroptosis, there is uncertainty surrounding the specificity of these transcription factors. Therefore, in-depth studies on both the specificity and
preclinical investigations are urgently needed. POST-TRANSLATIONAL MODIFICATIONS IN FERROPTOSIS Ferroptosis plays an important role in the regulation of ubiquitination, phosphorylation,
methylation, and acetylation. The key regulatory genes of ferroptosis, such as SLC7A11, GPX4, and voltage-dependent anion-selective channels (VDACs) [1,2,3], can be regulated by a series of
deubiquitinases, including ubiquitin-specific protease (USP) 11 [93], USP14 [94], and OTU domain-containing ubiquitin aldehyde-binding protein 1 [95], as well as ubiquitinases such as neural
precursor cell expressed developmentally downregulated protein 4 (NEDD4) [96], NEDD4 ligase [17], and so on. Among the ubiquitinases, NEDD4 is predicted to be the major E3 ligase mediating
VDAC1 degradation in melanoma. Another study has confirmed that endogenous VDAC1 interacts with NEDD4, and this contact was enhanced by erastin treatment. However, deletion of VDAC2/3
prevented erastin-induced ferroptosis. Interestingly, these effects were amplified following NEDD4 silencing. Additionally, an in-depth study has revealed that K63, K90, and K163 of the VDAC
subtype are critical for NEDD4-mediated ubiquitination [96]. Moreover, ferroptosis activity can be regulated by direct phosphorylation of ACSL4 or associated phosphorylation of SLC7A11.
Specifically, protein kinase C βII accelerates ferroptosis by directly phosphorylating ACSL4 at Thr328, which enhances the efficacy of immunotherapy in patients with melanoma [97]. In a
preclinical study, phosphorylation of beclin 1 at S90/93/96 contributed to its complexation with SLC7A11 and subsequent lipid peroxidation in ferroptosis, thereby prolonging the survival of
mice with pancreatic cancer [98]. In line with the above studies, numerous genes have been reported to regulate ferroptosis by targeting the SLC7A11/GPX4 axis through methylation.
Clinically, the high expression of GPX4 may be related to the low level of DNA methylation in multiple cancers. Furthermore, histone H3 lysine-4 trimethylation and acetylation of histone H3
on lysine 27 have been demonstrated to be enriched at the upstream site of GPX4 in different types of cancer tissues, indicating that the high expression of GPX4 may be the result of
methylation [99]. An additional study has evaluated the changes in total DNA methylation levels in the multiple myeloma cell lines MM1S and MM1R treated with the GPX4 inhibitor RSL3 and has
demonstrated that both MM1S and MM1R converge toward a similar methylation profile under ferroptosis conditions [100]. Besides, a previous study from the Hasegawa group has reported that the
ferroptosis inhibitor sulfasalazine, which targets SLC7A11, can increase DNA methylation on the mucin 1 gene (_MUC1_) promoter to regulate _MUC1_ gene transcription in triple-negative
breast cancer [101]. Moreover, acetylation also has been reported to be involved in the regulation of ferroptosis. For example, Jiang et al. have demonstrated that p533KR (K117/161/162)
regulates SLC7A11 expression and induces ferroptosis [78]. More interestingly, the loss of acetylation at K98 does not affect the transcriptional activity of p53. However, combining the
P533KR and K98 mutations renders P53 utterly incapable of regulating SLC7A11, indicating that K98 acetylation of p53 plays a key role in the inhibition of _SLC7A11_ expression and
p53-mediated ferroptosis [102]. Nevertheless, regulation of ferroptosis by acetylation requires more research, and the specific mechanism needs to be further elucidated. Therefore, the
identification of additional regulators of ferroptosis and acetylation will be necessary for clarifying their roles in cancer. FERROPTOSIS PLAYS A DUAL ROLE IN CANCER FERROPTOSIS AS A TUMOR
SUPPRESSOR EVENT Accumulating evidence indicates that ferroptosis acts as a tumor suppressor and influences the cycle, proliferation, and progression of cancer. So far, in addition to
blood-related cancers [103, 104], ferroptosis also acts as a tumor suppressor in multiple solid tumors, including lung cancer [105], breast cancer [106], pancreatic cancer [107], colorectal
cancer [108], liver cancer [109], esophageal squamous cell carcinoma [110], gastric cancer [111], and melanoma [112]. Moreover, previous studies have revealed that cytosolic aspartate
aminotransaminase (GOT1) is essential for redox balance and the proliferation of pancreatic ductal adenocarcinoma [113]. Recently, suppression of GOT1 has been demonstrated to inhibit GSH
biosynthesis by increasing GSSG and NADP+, which enhance the effect of butionine sulfoximine (BSO) on cancer cell proliferation. Furthermore, the simultaneous administration of GOT1 and BSO
has been shown to significantly slow the progression of xenograft tumors, and GOT1 combined with RSL3 significantly inhibits cell proliferation and enhances cytotoxicity. Interestingly,
inhibition of GOT1 combined with RSL3 or erastin can increase lipid ROS accumulation, and this enhancement can be reversed by cotreatment with the lipophilic antioxidant ferrostatin-1 [114].
On the other hand, ferroptosis can cause the migration and invasion of cancer cells to be inhibited. For example, it has been demonstrated that Krüppel-like factor 2 regulates ferroptosis
through GPX4, hence preventing cancer cell migration and invasion in clear cell renal cell carcinoma [115]. Another study has revealed that SLC7A11 promotes the migration and invasion of
renal cancer cells by enhancing GPX4 export [116]. However, in colorectal cancer cells, GPX4 can be negatively regulated by acyl-coenzyme A dehydrogenase, short/branched chain (ACADSB). The
overexpression of ACADSB reduces the migration and invasion of SW620 and LoVo cells by increasing intracellular Fe2+ and decreasing the expression of GPX4 and glutathione reductase [117]. In
addition to GPX4 and SLC7A11, another key regulator of ferroptosis, ACSL4, has been reported to act as a direct downstream target of miR-211-5p in HCC, and miR-211-5p suppresses malignant
phenotypes such as cell proliferation, migration, and invasion by inhibiting ACSL4 expression [118]. Along with ferroptosis blocking cell proliferation, migration, and invasion, an
investigation from the Feng group has revealed that ferroptosis is negatively associated with metastasis [119]. They found that lymph node metastases were more common in SLC7A11-positive
patients than in SLC7A11-negtive patients, suggesting that SLC7A11 expression is linked to tumor development and metastasis in esophageal squamous cell carcinoma [119]. Furthermore, SLC7A11
overexpression in human HCC is strongly linked with worse tumor differentiation, a higher tumor-nodule-metastasis stage, as well as a poor prognosis [120]. Additionally, Liu et al. have
demonstrated that ferroptosis resistance increase the probability of metastasis and have further confirmed that 27-hydroxycholesterol (27HC) is an abundant circulating cholesterol metabolite
that can be primarily used to screen cells for increased lipid biosynthesis. The enhanced tumorigenic and metastatic activities of 27HC-resistant cells can be reversed by GPX4 inhibition
[121]. Similarly, neratinib, a potent irreversible pan-tyrosine kinase inhibitor, has been shown to promote ferroptosis and to inhibit brain metastasis in a novel syngeneic model of
spontaneous human epidermal growth factor receptor 2-positive breast cancer metastasis [122]. These data further support the role of ferroptosis in inhibiting tumor metastasis. Emerging
evidence has suggested that ferroptosis is an inhibitor of prognosis and overall survival in clinical. The most direct evidence is that SLC7A11 has been shown to be significantly elevated in
RCC. Specifically, as an independent predictive factor, SLC7A11 overexpression is associated with a poor clinical prognosis [116]. ATP-binding cassette subfamily C member 5 (ABCC5) also has
been found to be upregulated in HCC clinical samples and negatively correlated with ferroptosis by increasing the intracellular GSH levels and stabilizing SLC7A11. More importantly, HCC
cells resistant to sorafenib have shown a substantial increase in ABCC5 expression, which is strongly associated with a worse prognosis [123, 124]. In addition, it has reported been reported
that ACSL4 and GPX4 regulate ferroptosis in opposite ways. Unlike GPX4, which has been demonstrated to be downregulated in breast cancer cells, ACSL4 has been verified to be substantially
expressed in breast cancer tissues [125, 126]. Moreover, ACSL4 and GPX4 expression, alone or combined, can be used as an independent prognostic factor for disease-free survival. Patients
with high ACSL4 expression have a better overall survival, while a higher GPX4 expression is associated with a better distant metastasis-free survival [127]. Taken together, the above
studies suggest that ferroptosis can act as a tumor suppressor through different molecular and cellular mechanisms. FERROPTOSIS AS A TUMOR ACTIVATOR Previous studies have indicated that GPX4
inactivation and ferroptosis caused by iron accumulation are important factors in tumor suppression. However, recent studies suggest that ferroptosis may function as a potential carcinogen
under special conditions. For example, knockdown of _Gpx4_ or a high-iron diet significantly increased the pancreatic weight, pancreatic intraepithelial neoplasia formation, and stromal
response as well as increased the mortality in Kirsten rat sarcoma viral oncogene homolog (Kras)4-driven animals. Nevertheless, the number of typical acinar cells was drastically diminished.
More importantly, tumor metastases to the liver and lungs were most prevalent in 10–12-month Pdx1-Cre, KrasG12D/+, Gpx4−/− mice. These results suggest that the lack of _Gpx4_ or a high-iron
diet speeds up the development of pancreatic ductal adenocarcinoma induced by _Kras_ [128]. Moreover, Ma et al. have shown that CD36 facilitates fatty acid absorption via tumor-infiltrating
CD8+ T cells. In particular, the upregulation of CD36 reduces the antitumor activity of CD8+ T cells by inducing lipid peroxidation and ferroptosis. Of note, the antitumor ability of CD8+ T
cells can be obviously renewed with CD36 deletion or suppression of ferroptosis [129]. However, the underlying regulatory mechanisms and specific pathways remain largely unknown. THE
PROSPECT OF FERROPTOSIS IN CANCER THERAPY Although ferroptosis plays a dual role in the development of tumors, it is typically considered as a well-known tumor suppressor in specific
situations, as mentioned above. Due to the fact that ferroptosis inducers can be synthesized and have been verified in multiple in vivo experiments. Ferroptosis inducers have the potential
to represent an anticancer therapy for clinical application after large-scale population validation. In line with this notion, numerous studies have developed different inducers to activate
ferroptosis and explored the role of ferroptosis in different cancer therapies, such as such as chemotherapy, immunotherapy, radiotherapy, nanotherapy, SDT and PDT. ROLE OF FERROPTOSIS IN
CHEMOTHERAPY Chemotherapy is one of the main methods to treat cancer. Despite the clinical success of cancer chemotherapy, drug resistance continues to be a major and complex issue.
Fortunately, some synthetic and natural compounds have demonstrated promising effects by activating ferroptosis, effectively inhibiting acquired tumor drug resistance and optimizing cancer
efficacy (summarized in Table 3). SYNTHETIC COMPOUNDS At present, the classic activators of ferroptosis mainly include sorafenib and cisplatin, which have been used clinically, and some
preclinical experimental compounds such as erastin and RSL3. Among them, sorafenib is the first drug approved for the treatment of HCC and RCC. However, recent research regarding pancreatic
adenocarcinoma and colon carcinoma have shown that the anticancer activity of sorafenib mainly depends on the inhibition of SLC7A11/GPX4 activity to induce ferroptosis; the activation of
ferroptosis in turn promotes the anticancer effect of sorafenib [124, 130, 131]. On the other hand, cisplatin, one of the best metal-based chemotherapeutic drugs [132], has been demonstrated
to play a non-negligible role in the regulation of lung cancer progression via targeting ferroptosis. Mechanistically, invasion and migration assays have shown that the combination of
cisplatin and RSL3 can suppress the invasion and migration of H1299 and A549 cells more effectively. Moreover, inhibition of GPX4 by RSL3 has been reported to markedly enhance the anticancer
effects of cisplatin by inhibiting tumor growth [99]. In addition, the activity of erastin and RSL3 has been used as a single or combined therapy for prostate cancer [133]. Ferroptosis
induced by erastin and RSL3 not only inhibits cell migration and invasion, but it also delays the growth of drug-resistant prostate cancer tumors. It is worth noting that this treatment did
not cause side effects such as weight loss or pain in mice [133]. In view of the complex antitumor effect of ferroptosis, more types of ferroptosis inducers have been actively developed. For
example, the selective small molecule inhibitor, (+)-JQ1(JQ1) is the first potent and selective inhibitor of the bromodomain-containing protein 4 signaling pathway that has been reported to
inhibit the proliferation of esophageal squamous cell carcinoma [134] and multiple myeloma [135]. In addition, Sui et al. have revealed that JQ1 induces ferroptosis by increasing ROS levels
in breast cancer and lung squamous cell carcinoma cells. The rate of cell death was dramatically accelerated when JQ1 was coupled with the ferroptosis inducer RSL3. Further experiments have
confirmed that the anticancer effect of JQ1 in breast cancer and lung squamous cell carcinoma cells can be enhanced by ferroptosis inducers [136]. Additionally, significant decreases in the
tumor volume, size, and weight were observed after treatment with the broad spectrum deubiquitinase inhibitor palladium pyrithione complex (PdPT), and downregulation of GPX4 levels was
detected, implying that PdPT may exert tumor-suppressing effects by relying on ferroptosis [137]. In line with these findings, the local anesthetic lidocaine has been proven to suppress the
mRNA expression of SLC7A11 and to promote ferroptosis in ovarian and breast cancer cells in a dose-dependent manner, thereby repressing tumor growth [138]. Similarly, in bladder cancer, it
has been shown that another widely used local anesthetic, bupivacaine, can inhibit the expression of SLC7A11 and GPX4 but increase the levels of Fe2+ and ROS, thus triggering ferroptosis and
restraining the growth of xenografted tumors [139]. Furthermore, ketamine has been studied as rapid-acting anesthetic in cancer treatment [140]. A recent medical study has validated that
ketamine can induce ferroptosis and limit cell proliferation, and its effect can be reversed by GPX4 overexpression in liver cancer [67]. Strikingly, emerging evidence has suggested that the
small molecule MMRi62 can act as a novel ferroptosis inducer in pancreatic cancer; its use restricts tumor growth and metastasis by promoting the degradation of mutant p53 [141]. These
findings lay the foundation for the clinical application of novel ferroptosis inducers and also encouraged us to explore whether additional ferroptosis inducers can cure cancer in an
independent way. It would also be very interesting to test the combined effects of ferroptosis inducers with other existing anticancer drugs. NATURAL COMPOUNDS Compared to traditional cancer
therapies and drugs, natural compounds have shown unique advantages. Most importantly, they have attracted more and more attention because of their multiple benefits with minimal side
effects [142]. Several studies have shown that 6-gingerol, a natural phenol in ginger (_Zingiber officinale_ Roscoe), has potent anti-inflammatory, antitumor, and antioxidant effects
[143,144,145]. It also has been demonstrated that 6-gingerol can significantly increase ROS and iron concentrations in vitro and in vivo, decrease the survival and proliferation of lung
cancer cells, and reduce the tumor volume and weight in nude mice [94]. In triple-negative breast cancer, a derivative of the natural product parthenolide (DMOCPTL) has been reported to
increase ROS accumulation in a time- and dose-dependent manner by directly targeting GPX4. Of note, DMOCPTL can significantly inhibit tumor growth and prolong the survival of mice, and it
has no obvious toxicity [146]. Further research has demonstrated that a bufadienolide called BT, which can be isolated from the plant _Venenum bufonis_, exerts a similar effect.
Specifically, BT has been shown to increase intracellular Fe2+ in vitro. In a xenograft model, BT inhibited tumor growth without significantly changing the mouse weight, downregulated GPX4,
and induced lipid peroxidation, further supporting that BT exerts a tumor-suppressing effect with fewer side effects [147]. Besides, a recent study has shown that the natural coumarin
alloimperatorin, found in the extract of _Angelica dahurica_, has anticancer properties [148]. Functionally, alloimperatorin directly decreases the expression levels of SLC7A11 and GPX4,
thereby inhibiting the growth and invasion of breast cancer cells [149]. However, to date, there is still no direct clinical evidence of its therapeutic effect on human cancer. Therefore,
in-depth research is required to determine whether these natural extracts have broad-spectrum effects, few side effects, and therapeutic effectiveness against human cancer. ROLE OF
FERROPTOSIS IN RADIOTHERAPY Radiotherapy leads to the accumulation of ROS via the radiolysis of cellular water, which damages biomolecules including lipids; therefore, it is speculated that
radiotherapy may be related to ferroptosis. To explore this possibility, Lang et al. performed staining with C11BODIPY, a lipophilic redox-sensitive dye, in HT1080 cells and found that
radiotherapy resulted in an increase in lipid ROS concentrations. Furthermore, deletion of the ferroptosis-promoting gene _ACSL4_ reduced the efficacy of radiotherapy. Most importantly,
ferroptosis-resistant tumors remained radioresistant in vivo even at higher concentrations of radiotherapy [150]. These effects also have been confirmed in subsequent experiments. For
example, data from The Cancer Genome Atlas have revealed that _SLC7A11_ plays a role in the radioresistance of gliomas [151]. Additional experiments have demonstrated that the systemic XC—
inhibitor imidazole ketone erastin and RSL3 enhance the radiosensitivity of cancer cells through lipid peroxidation. The combined treatment with imidazole ketone erastin and sorafenib also
has been reported to promote the inhibitory effect of radiation on tumor growth in a xenografted mouse model of sarcoma [151]. In line with these findings, Lei et al. have shown that
ionizing radiation (IR) not only induces ROS but also increases the expression of ACSL4. In addition, inhibition of ACSL4 has been demonstrated to largely abolish IR-induced ferroptosis and
to enhance radio resistance. Notably, IR increases the expression of SLC7A11 and promotes radio resistance by suppressing ferroptosis. Moreover, the inhibition of SLC7A11 or GPX4 has been
shown to enhance IR sensitivity in radioresistant cancer cells and xenografts [152]. Further studies have found that radiotherapy induces ferroptosis, which in turn is associated with a
better response to radiotherapy and a longer survival in esophageal cancer patients [152]. Collectively, ferroptosis plays a crucial role in radiotherapy-mediated tumor suppression; thus,
inducing ferroptosis in radiotherapy-resistant tumors is a promising strategy for radio sensitization. However, for ferroptosis-resistant tumors that may also be radioresistant, ways to
minimize their dual drug resistance must be discovered. ROLE OF FERROPTOSIS IN IMMUNOTHERAPY Immunotherapy is a new treatment method following surgery, radiotherapy, and chemotherapy, which
brings the possibility of eradicating cancer. However, the immune escape is a major factor leading to the poor efficacy of tumor immunotherapy and has become a major obstacle of this method.
Therefore, blocking the immune escape is one of the key issues to improve the effect of tumor immunotherapy. Ferroptosis has been reported to suppress tumor growth and metastasis via immune
checkpoints. Previous studies have predicted that the immune checkpoint molecule programmed death-1 is positively correlated with the expression level of ACSL4 but negatively correlated
with the expression levels of GPX4 and HSPB1 in clear cell renal cell carcinoma [153]. In line with this possibility, Liao et al. have demonstrated an increased overall survival or
progression-free survival in patients with high ACSL4 expression following immune checkpoint blockade therapy [154]. Further experiments have indicated that BNP@R (including RSL3,
PEG-_b_-P(DPA-_r_-PPa), and phenylboronic acid moieties grafted onto the mPEG-_b_-P (DPA-_r_-GC backbone)) activate the T-cell immune response by inducing ferroptosis in melanoma. Treatment
combined with BNP@R and GPX4 inhibition has been shown to significantly induce programmed death-ligand 1 (PD-L1) expression both in vitro and in vivo. More importantly, combination treatment
of BNP@R + laser and anti-PD-L1 induces ROS accumulation, which in turn significantly suppresses tumor growth and prolongs survival in mice [155]. More convincing evidence is that PD-L1
blockade treatment directly led to an increase of ROS in CD45-ID8 cells and effectively reduced tumor growth [156]. These data further support that tumor immune checkpoint inhibitor therapy
based on ferroptosis is expected to provide a new strategy for tumor immunotherapy. Besides, recent studies also have shown that ferroptosis can inhibit tumor cell immune escape via
activating tumor immune cells, such as macrophages, natural killer cells, and T cells. To explore the effect of GPX4 on T cell physiological responses, Matsushita et al. constructed T
cell-specific _Gpx4_-deficient mice and found that neither antigen-specific CD8+ nor CD4+ T cells lacking Gpx4 could expand and protect them from infection and that _Gpx4_-deficient T cells
rapidly accumulated membrane lipid peroxides [157]. Additional studies have elucidated that CD8+ T cell-derived interferon gamma increases the binding of signal transducer and activator of
transcription 1 (STAT1) to the transcription start site of SLC7A11 and inhibits its transcription. Thus, loss of STAT1 in tumor cells abrogates interferon gamma-mediated downregulation of
SLC7A11 while reversing RSL3-induced lipid peroxidation and ferroptosis [156]. Macrophages are important immune cells in the human body and are mainly divided into M1 macrophages (typically
activated or proinflammatory macrophages) and M2 macrophages (alternatively activated or anti-inflammatory macrophages) [158]. In general, M1 macrophages can generate ROS through the NADPH
oxidase 2 pathway to help kill pathogens during immunity. Advanced glycation end products can induce M1 polarization in macrophages by stimulating the ROS/toll-like receptor 4/STAT1 pathway
[159]. Moreover, iron overload-induced ROS generation and p53 acetylation also contribute to M1 polarization [160]. However, the levels of M1 markers such as interleukin-6, tumor necrosis
factor-alpha, and interleukin-1beta can be increased by iron overload, while the levels of M2 markers are suppressed, thereby promoting the polarization of M1 macrophages [161]. This opens
the door to the possibility of using M2 repolarization of tumor-associated macrophages as a method of tumor immunotherapy. Notably, it has been recently shown that M2 macrophages transform
into M1 macrophages through the ferroptosis pathway, which can enhance anti-programmed cell death protein 1 immunotherapy in liver cancer [162]. In addition, bioinformatics analysis has
revealed that high ferroptosis risk levels are closely associated with natural killer cell inactivation in breast cancer [163]. Moreover, in-depth research has determined that CAR-NK92MI
cells can kill prostate cancer cells by inducing ferroptosis [164]. Collectively, ferroptosis has shown much promise in immunotherapy, and these findings have been confirmed in vivo and in
vitro. Nonetheless, further prospective clinical trials are needed to validate its safety and efficacy as well as its precise mechanism. ROLE OF FERROPTOSIS IN NANOTHERAPY Targeted drug
delivery based on nanocarriers has the potential to improve the utilization rate of drugs, increase the intracellular uptake capacity of drugs, and reduce the toxic and side effects of drugs
to a certain extent. Nanoparticles that are loaded with chemicals or biological components provide novel promise for enhancing the efficiency of existing inducers of ferroptosis and cancer
therapy. Specifically, combination therapy of poly(ethylene glycol)-coated silica nanoparticles and amino acid starvation synergistically induce ferroptosis, and high-dose particle delivery
can inhibit tumor growth and even lead to tumor regression [165]. Notably, the leakiness of the tumor vasculature is thought to be due to systemically injected nanoparticles accumulating in
tumor tissue while causing nutrient deprivation within the tumor [166]. Interestingly, tumor blood vessels and nutrients are not destroyed when nanoparticles induce ferroptosis [165], laying
the foundation for subsequent research. Another study has demonstrated that the antitumor activity of imidazole ketone erastin delivered in polyethylene glycol poly(lactic-co-glycolic acid)
nanoparticles was enhanced in a mouse lymphoma model [167]. More recently, Li et al. have engineered a dual catalytic nanomedicine named Lipo-ART@CPNs (co-loading CuO2 nanodots and
artemisinin) which can increase ROS levels, promote iron release, and lead to ferroptosis. Under the treatment of Lipo-ART@CPNs, the tumor inhibition rate of Lewis lung carcinoma-bearing
mice was found to be increased by 18% compared to the non- Lipo-ART@CPNs treatment groups (58%). Furthermore, the tumor cells were obviously destroyed [168]. Consistent with this finding,
copper nanodots (Cu NDs) that induce excess lipid peroxide and inhibit GPX4 and SLC7A11 to induce ferroptosis have been developed [169]. Similarly, in comparison to the control group, Cu NDs
treatment group increased tumor growth inhibition to 90.8% after ultrasound (US). Most importantly, there were no statistically significant weight changes among all groups of mice,
providing strong evidence for the safety and efficacy of this therapy [169]. Although further evidence is needed to support the superiority of this therapy, it will be interesting and
promising to explore whether synergistic therapies combining ferroptosis and nanotherapy would be a more effective cancer treatment strategy. ROLE OF FERROPTOSIS IN SONODYNAMIC THERAPY SDT
is a treatment method that associates US sensitizer, oxygen with low intensity US to trigger ROS accumulation to kill cancer cells. Emerging evidence showed that ferroptosis combined with
SDT has great clinical application potential due to its low cost, high penetration and non-invasive removal of solid tumors in a fixed way [170, 171]. Specifically, a previous report
revealed that a specific sensitizer including Fe/Mn component could trigger ferroptosis in breast cancer under US [170]. One reason is that it can catalyze Fenton reaction to produce ROS, as
well as increased the consumption of glutathione in cells. Of note, it can be clearly observed in mice that this treatment significantly inhibited the growth of tumor without affecting the
blood index and weight of mice, which proved that the combination of ferroptosis and SDT can produce anti-tumor effect with minimal side effects [170]. Similar effects have also been
verified in glioma. Zhu et al. developed a sensitizer called PIOC@CM NPs which combination of Fe3O4 and Ce6 was tested in vitro and in vivo [171]. The results showed that PIOC@CM NPs
produced excessive ROS after US, which could not only reduce the cytotoxicity, but also increase the level of ACSL4. It is noteworthy that the introduction of ferroptosis and SDT
significantly reduced the tumor volume and prolonged the survival of mice [171]. Further study constructed Aza-boron-dipyrromethene dyne (Aza-BDY, both an ferroptosis inducer and a
sensitizer) and found it reduced cell viability after US [172]. Surprisingly, the synergistic effect of ferroptosis and SDT made the tumor inhibition rate reach 97.5%, and even eliminated
the mice tumor completely [172]. Nevertheless, it is still unknown whether it can suppress recurrence and metastasis of cancer. Further in-depth research is required to determine whether
ferroptosis combined with SDT has broad-range effects in different cancer cells and clinical efficacy. ROLE OF FERROPTOSIS IN PHOTODYNAMIC THERAPY PDT is a non-invasive treatment method that
injects photosensitizer (PS) into the body, and then irradiates it at a specific wavelength to produce ROS, leading to oxidative stress and cell damage to treat tumor [173]. It exerts many
effects combating different cancers, such as esophageal cancer, lung cancer and skin cancer [174,175,176]. However, due to the presence of intracellular antioxidants, especially GSH can
effectively eliminate ROS, which seriously reduces the efficiency of PDT. To overcome this limitation, researchers introduced ferroptosis with PDT to enhance anti-tumor effect. A previous
study conducted by Zhu et al. combined PS chlorine e6 (Ce6) and ferroptosis inducer erastin to form Ce6-erastin complex and verified in oral tongue square cell carcinoma (OTSCC) mice.
Immunohistochemical analysis showed that the induction of PDT and ferroptosis decreased the expression of Ki-67 and significantly inhibited the expression of SLC7A11 compared with the
control group [173]. Subsequently, further research developed PS called AE@RBC /Fe NCs triggered ferroptosis by reducing GSH level. Surprisingly, the participation of PDT further accelerated
the consumption of GSH and increased the level of MDA. Of note, the tumor inhibition rate reached 95.8%, and the survival time of mice was significantly prolonged without affecting the
liver and kidney functions of oral squamous cell carcinoma (OSCC) mice [177]. Furthermore, another PS named Fe3O4@Lipid NPs was recently reported to increase ROS level in a time and dose
-dependent manner under irradiation in colorectal cancer. More importantly, it can be observed that the tumor was significantly necrotic, and the volume was significantly reduced without
reducing the weight of mice [178]. These results supported the conclusion that the combination of ferroptosis and PDT has the superiority of anticancer effect. However, it remains to be
explored whether this integration therapy has specificity and wide applicability in multiple cancers in terms of time, dose and irradiation wavelength. CONCLUSIONS AND PERSPECTIVES Since its
discovery in 2012, the role of ferroptosis in regulating a variety of cellular processes and different diseases, especially cancer, has been extensively studied [179, 180]. Ferroptosis
plays a dual role in human tumorigenesis due to the complicated tumor microenvironment. Therefore, considering the tumor-suppressing effect of ferroptosis, the development of more specific
inducers of ferroptosis may be a potential and promising cancer treatment strategy. In particular, due to the fact that various cancer cells have different sensitivities to ferroptosis,
determining which cancers are more amenable to treatments incorporating ferroptosis will be an area of active research in the next few years. To date, only some classical compounds such as
erastin, RSL3, etc. are more specific for ferroptosis, while other inducers, including sorafenib (the first line drug in unresectable or advanced HCC and RCC), are not specific for
ferroptosis [81, 181]. With this concept in mind, it is necessary and urgent to screen and develop more specific activators of ferroptosis. On the other hand, using natural compounds or
nanoparticles as ferroptosis inducers may be a safe and effective cancer treatment strategy due to their properties and few side effects. More importantly, combine ferroptosis inducers with
other anticancer therapies will provide new sights for cancer treatment. With the exception of directly targeting ferroptosis, other approaches should be explored, such as the induction of
ferroptosis through modulation of ncRNAs, transcription factors, and post-translational modifications (as summarized in Table 2). Given that ferroptosis is a double-edged sword in
tumorigenesis, it will be interesting to explore the physiological role of ferroptosis in the progression of various cancers through conditional knockout or knockin mouse models. Going
forward, cancer type-specific animal models of ferroptosis will facilitate and improve the development of specific ferroptosis promoters, and large-scale clinical trials will also help to
accelerate their clinical translation. In the near future, it is believed that ferroptosis inducers with an optimal specificity and efficacy will be developed and used to treat various types
of cancer. DATA AVAILABILITY All data in this study are available. REFERENCES * Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020:
GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71:209–49. Article PubMed Google Scholar * Mun EJ, Babiker HM, Weinberg U,
Kirson ED, Von Hoff DD. Tumor Treating Fields: A Fourth Modality in Cancer Treatment. Clin Cancer Res. 2018;24:266–75. Article CAS PubMed Google Scholar * Dixon SJ, Lemberg KM, Lamprecht
MR, Skouta R, Zaitsev EM, Gleason CE, et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell. 2012;149:1060–72. Article CAS PubMed PubMed Central Google Scholar *
Dixon SJ, Winter GE, Musavi LS, Lee ED, Snijder B, Rebsamen M, et al. Human Haploid Cell Genetics Reveals Roles for Lipid Metabolism Genes in Nonapoptotic Cell Death. ACS Chem Biol.
2015;10:1604–9. Article CAS PubMed PubMed Central Google Scholar * Doll S, Freitas FP, Shah R, Aldrovandi M, Silva MC, Ingold I, et al. FSP1 is a glutathione-independent ferroptosis
suppressor. Nature. 2019;575:693–8. Article CAS PubMed Google Scholar * Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35:495–516. Article CAS PubMed
PubMed Central Google Scholar * Ferrer CC, Berthenet K. Apoptosis – Fueling the oncogenic fire. FEBS J. 2021;288:4445–63. Article Google Scholar * Klionsky DJ, Petroni G, Amaravadi RK,
Baehrecke EH, Ballabio A, Boya P, et al. Autophagy in major human diseases. EMBO J. 2021;40:e108863. Article CAS PubMed PubMed Central Google Scholar * Onorati AV, Dyczynski M, Ojha R,
Amaravadi RK. Targeting Autophagy in Cancer. Cance.r 2018;124:3307–18. Google Scholar * Cobine PA, Brady DC. Cuproptosis: Cellular and molecular mechanisms underlying copper-induced cell
death. Mol Cell. 2022;82:1786–7. Article CAS PubMed Google Scholar * Tsvetkov P, Coy S, Petrova B, Dreishpoon M, Verma A, Abdusamad M, et al. Copper induces cell death by targeting
lipoylated TCA cycle proteins. Science. 2022;1261:1254–61. Article Google Scholar * Gong Y, Fan Z, Luo G, Yang C, Huang Q, Fan K, et al. The role of necroptosis in cancer biology and
therapy. Mol Cancer. 2019;18:1–17. Article CAS Google Scholar * Frank D, Vince JE. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ.
2019;26:99–114. Article PubMed Google Scholar * Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021;6:128. Article
PubMed PubMed Central Google Scholar * Tang D, Chen X, Kroemer G. Cuproptosis: a copper-triggered modality of mitochondrial cell death. Cell Res. 2022;35:417–8. Article Google Scholar *
Ingold I, Berndt C, Schmitt S, Doll S, Poschmann G, Buday K, et al. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis. Cell. 2018;172:409–22. Article
CAS PubMed Google Scholar * Wang Y, Liu Y, Liu J, Kang R, Tang D. NEDD4L-mediated LTF protein degradation limits ferroptosis. Biochem Biophys Res Commun. 2020;531:581–7. Article CAS
PubMed Google Scholar * Liu J, Kang R, Tang D. Signaling pathways and defense mechanisms of ferroptosis. FEBS J. 2022;289:7038–50. Article CAS PubMed Google Scholar * Stockwell BR,
Jiang X, Gu W. Emerging Mechanisms and Disease Relevance of Ferroptosis. Trends Cell Biol. 2020;30:478–90. Article CAS PubMed PubMed Central Google Scholar * Yang Y, Zhu T, Wang X,
Xiong F, Hu Z, Qiao X, et al. ACSL3 and ACSL4, Distinct Roles in Ferroptosis and Cancers. Cancers (Basel). 2022;14:5896. Article CAS PubMed Google Scholar * Li Z, Chen L, Chen C, Zhou Y,
Hu D, Yang J, et al. Targeting ferroptosis in breast cancer. Biomark Res. 2020;8:58. Article PubMed PubMed Central Google Scholar * Zhao L, Zhou X, Xie F, Zhang L, Yan H, Huang J, et
al. Ferroptosis in cancer and cancer immunotherapy. Cancer Commun (Lond). 2022;42:88–116. Article PubMed PubMed Central Google Scholar * Wohlhieter CA, Richards AL, Uddin F, Hulton CH,
Quintanal-Villalonga À, Martin A, et al. Concurrent Mutations in STK11 and KEAP1 Promote Ferroptosis Protection and SCD1 Dependence in Lung Cancer. Cell Rep. 2020;33:108444. Article CAS
PubMed PubMed Central Google Scholar * Tang D, Chen X, Kang R, Kroemer G. Ferroptosis: molecular mechanisms and health implications. Cell Res. 2021;31:107–25. Article CAS PubMed Google
Scholar * Tsoi J, Robert L, Paraiso K, Galvan C, Sheu KM, Lay J, et al. Multi-stage Differentiation Defines Melanoma Subtypes with Differential Vulnerability to Drug-Induced Iron-Dependent
Oxidative Stress. Cancer Cell. 2018;33:890–904.e5. Article CAS PubMed PubMed Central Google Scholar * Viswanathan VS, Ryan MJ, Dhruv HD, Gill S, Eichhoff OM, Seashore-ludlow B, et al.
Dependency of a therapy_resistant state of cancer cells on a lipid peroxidase pathway. Nature. 2017;547:453–7. Article CAS PubMed PubMed Central Google Scholar * Tabnak P,
Hajiesmailpoor Z, Soraneh S. Ferroptosis in lung cancer: from molecular mechanisms to prognostic and therapeutic opportunities. Front Oncol. 2021;11:792827. Article CAS PubMed PubMed
Central Google Scholar * Wang D, Tang L, Zhang Y, Ge G, Jiang X, Mo Y, et al. Regulatory pathways and drugs associated with ferroptosis in tumors. Cell Death Dis. 2022;13:544. Article
PubMed PubMed Central Google Scholar * Bedard K, Krause K. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol Rev. 2007;87:245–313. Article CAS
PubMed Google Scholar * Cheng Y, Song Y, Chen H, Li Q, Gao Y, Lu G. Ferroptosis Mediated by Lipid Reactive Oxygen Species: A Possible Causal Link of Neuroinflammation to Neurological
Disorders. Oxid Med Cell Longev. 2021;2021:5005136. Article PubMed PubMed Central Google Scholar * Kirtonia A, Sethi G, Garg M, Artesunate ART. The multifaceted role of reactive oxygen
species in tumorigenesis. Cell Mol Life Sci. 2020;77:4459–83. Article CAS PubMed Google Scholar * Das UN. Saturated Fatty Acids, MUFAs and PUFAs Regulate Ferroptosis. Cell Chem Biol.
2019;26:309–11. Article CAS PubMed Google Scholar * Huang W, Aabed N, Shah YM, Physiology I, Arbor A, Arbor A, et al. Reactive oxygen species and ferroptosis at the nexus of inflammation
and colon cancer. Antioxid Redox Signal. 2023. https://doi.org/10.1089/ars.2023.0246. online ahead of print. * Su LJ, Zhang JH, Gomez H, Murugan R, Hong X, Xu D, et al. Reactive oxygen
species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid Med Cell Longev. 2019;2019:5080843. Article PubMed PubMed Central Google Scholar * Xu Y, Liu SY, Zeng L,
Ma H, Zhang Y, Yang H, et al. An enzyme-engineered nonporous copper(I) coordination polymer nanoplatform for cuproptosis-based synergistic cancer therapy. Adv Mater. 2022;34:e2204733.
Article PubMed Google Scholar * Bertheloot D, Latz E, Franklin BS. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cell Mol Immunol. 2021;18:1106–21. Article CAS
PubMed PubMed Central Google Scholar * Chen X, Comish PB, Tang D, Kang R. Characteristics and biomarkers of ferroptosis. Front Cell Dev Biol. 2021;9:637162. Article PubMed PubMed
Central Google Scholar * Bebber CM, Müller F, Clemente LP, Weber J, von Karstedt S. Ferroptosis in cancer cell biology. Cancers (Basel). 2020;12:164. Article CAS PubMed Google Scholar
* Santavanond JP, Rutter SF, Atkin-Smith GK, Poon IKH. Apoptotic bodies: mechanism of formation, isolation and functional relevance. Subcell Biochem. 2021;97:61–88. Article CAS PubMed
Google Scholar * Man SM, Karki R, Kanneganti TD. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol Rev.
2017;277:61–75. Article CAS PubMed PubMed Central Google Scholar * Yan N, Zhang JJ. Iron Metabolism, Ferroptosis, and the links with Alzheimer’s disease. Front Neurosci. 2020;13:1443.
Article PubMed PubMed Central Google Scholar * Ward RJ, Zucca FA, Duyn JH, Crichton RR, Zecca L. The role of iron in brain ageing and neurodegenerative disorders. Lancet Neurol.
2014;13:1045–60. Article CAS PubMed PubMed Central Google Scholar * Wang Y, Yu L, Ding J, Chen Y. Iron metabolism in cancer. Int J Mol Sci. 2019;20:95. Article Google Scholar * Ng SW,
Norwitz SG, Norwitz ER. The impact of iron overload and ferroptosis on reproductive disorders in humans: Implications for preeclampsia. Int J Mol Sci. 2019;20:3283. Article CAS PubMed
PubMed Central Google Scholar * Zhang C, Zhang F. Iron homeostasis and tumorigenesis: molecular mechanisms and therapeutic opportunities. Protein Cell. 2015;6:88–100. Article CAS PubMed
Google Scholar * Brookes MJ, Hughes S, Turner FE, Reynolds G, Sharma N, Ismail T, et al. Modulation of iron transport proteins in human colorectal carcinogenesis. Gut. 2006;55:1449–60.
Article CAS PubMed PubMed Central Google Scholar * Kuang Y, Wang Q. Iron and lung cancer. Cancer Lett. 2019;464:56–61. Article CAS PubMed Google Scholar * Li D, Li Y. The
interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct Target Ther. 2020;5:108. Article PubMed PubMed Central Google Scholar * Liang D, Minikes AM, Jiang X.
Ferroptosis at the intersection of lipid metabolism and cellular signaling. Mol Cell. 2022;82:2215–27. Article CAS PubMed PubMed Central Google Scholar * Lee J, Kim WK, Bae K, Lee SC,
Lee E. Lipid metabolism and ferroptosis. Biol (Basel). 2021;10:184. CAS Google Scholar * Kagan VE, Mao G, Qu F, Angeli JPF, Doll S, Croix CS, et al. Oxidized arachidonic and adrenic PEs
navigate cells to ferroptosis. Nat Chem Biol. 2017;13:81–90. Article CAS PubMed Google Scholar * Magtanong L, Ko PJ, To M, Cao JY, Forcina GC, Tarangelo A, et al. Exogenous
monounsaturated fatty acids promote a ferroptosis-resistant cell state. Cell Chem Biol. 2019;26:420–432.e9. Article CAS PubMed PubMed Central Google Scholar * Ubellacker JM, Tasdogan A,
Ramesh V, Shen B, Mitchell EC, Martin-Sandoval MS, et al. Lymph protects metastasizing melanoma cells from ferroptosis. Nature. 2020;585:113–8. Article CAS PubMed PubMed Central Google
Scholar * Hoy AJ. Tumour fatty acid metabolism in the context of therapy resistance and obesity. Nat Rev Cancer. 2021;21:753–66. Article CAS PubMed Google Scholar * Vettore L, Westbrook
RL, Tennant DA. New aspects of amino acid metabolism in cancer. Br J Cancer. 2019;122:150–6. Article PubMed PubMed Central Google Scholar * Dixon SJ, Patel D, Welsch M, Skouta R, Lee E,
Hayano M, et al. Pharmacological inhibition of cystine-glutamate exchange induces endoplasmic reticulum stress and ferroptosis. Elife. 2014;2014:e02523. Article Google Scholar * Wang X,
Wang Z, Cao J, Dong Y, Chen Y. Ferroptosis mechanisms involved in hippocampal-related diseases. Int J Mol Sci. 2021;22:9902. Article CAS PubMed PubMed Central Google Scholar * Koppula
P, Zhuang L, Gan B. Cystine transporter SLC7A11/xCT in cancer: ferroptosis, nutrient dependency, and cancer therapy. Protein Cell. 2021;12:599–620. Article CAS PubMed Google Scholar *
Forcina GC, Dixon SJ. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics 2019;19:e1800311. Article PubMed Google Scholar * Liu M. System Xc −: a key regulatory target
of ferroptosis in cancer. Invest N. Drugs. 2021;39:1123–31. Article CAS Google Scholar * Xu P, Wang Y, Deng Z, Tan Z, Pei X. MicroRNA-15a promotes prostate cancer cell ferroptosis by
inhibiting GPX4 expression. Oncol Lett. 2022;23:67. Article CAS PubMed PubMed Central Google Scholar * Liu L, Yao H, Zhou X, Chen J, Chen G, Shi X, et al. MiR-15a-3p regulates
ferroptosis via targeting glutathione peroxidase GPX4 in colorectal cancer. Mol Carcinog. 2022;61:301–10. Article CAS PubMed Google Scholar * Gomaa A, Peng D, Chen Z, Soutto M, Abouelezz
K, Corvalan A, et al. Epigenetic regulation of AURKA by miR-4715-3p in upper gastrointestinal cancers. Sci Rep. 2019;9:16970. Article PubMed PubMed Central Google Scholar * Deng S, Wu
D, Li L, Liu T, Zhang T, Li J, et al. miR-324-3p reverses cisplatin resistance by inducing GPX4-mediated ferroptosis in lung adenocarcinoma cell line A549. Biochem Biophys Res Commun.
2021;16:54–60. Article Google Scholar * Wei D, Ke YQ, Duan P, Zhou L, Wang CY, Cao P. MicroRNA-302a-3p induces ferroptosis of non-small cell lung cancer cells via targeting ferroportin.
Free Radic Res. 2021;55:821–30. Article CAS PubMed Google Scholar * Bai T, Liang R, Zhu R, Wang W, Zhou L, Sun Y. MicroRNA-214-3p enhances erastin-induced ferroptosis by targeting ATF4
in hepatoma cells. J Cell Physiol. 2020;235:5637–48. Article CAS PubMed Google Scholar * He GN, Bao NR, Wang S, Xi M, Zhang TH, Chen FS. Ketamine induces ferroptosis of liver cancer
cells by targeting lncRNA PVT1/miR-214-3p/GPX4. Drug Des Devel Ther. 2021;15:3965–78. Article PubMed PubMed Central Google Scholar * Li YZ, Zhu HC, Du Y, Zhao HC, Wang L. Silencing
lncRNA SLC16A1-AS1 induced ferroptosis in renal cell carcinoma through miR-143-3p/SLC7A11 signaling. Technol Cancer Res Treat. 2022;21:15330338221077803. Article PubMed PubMed Central
Google Scholar * Wang Z, Chen X, Liu N, Shi Y, Liu Y, Ouyang L, et al. A nuclear long non-coding RNA LINC00618 accelerates ferroptosis in a manner dependent upon apoptosis. Mol Ther.
2021;29:263–74. Article CAS PubMed Google Scholar * Chen C, Zhao J, Liu JN, Sun C. Mechanism and Role of the Neuropeptide LGI1 Receptor ADAM23 in Regulating Biomarkers of Ferroptosis and
Progression of Esophageal Cancer. Dis Markers. 2021;2021:9227897. Article PubMed PubMed Central Google Scholar * Yu W, Invasion C, Hospital X, South C, Hospital SX, Program CE, et al. A
G3BP1-interacting lncRNA promotes ferroptosis and apoptosis in cancer via nuclear sequestration of p53. Cancer Res. 2018;78:3484–96. Article PubMed PubMed Central Google Scholar * Gai
C, Liu C, Wu X, Yu M, Zheng J, Zhang W, et al. MT1DP loaded by folate-modified liposomes sensitizes erastin-induced ferroptosis via regulating miR-365a-3p/NRF2 axis in non-small cell lung
cancer cells. Cell Death Dis. 2020;11:751. Article CAS PubMed PubMed Central Google Scholar * Jiang M, Mo R, Liu C, Wu H. Circ_0000190 sponges miR-382-5p to suppress cell proliferation
and motility and promote cell death by targeting ZNRF3 in gastric cancer. J Biochem. 2022;mvac003. https://doi.org/10.1093/jb/mvac003. online ahead of print. * Wang Y, Chen H, Wei X.
Circ_0007142 downregulates miR-874-3p-mediated GDPD5 on colorectal cancer cells. Eur J Clin Invest. 2021;51:e13541. Article CAS PubMed Google Scholar * Bazhabayi M, Qiu X, Li X, Yang A,
Wen W, Zhang X, et al. CircGFRA1 facilitates the malignant progression of HER-2-positive breast cancer via acting as a sponge of miR-1228 and enhancing AIFM2 expression. J Cell Mol Med.
2021;25:10248–56. Article CAS PubMed PubMed Central Google Scholar * Ou R, Lu S, Wang L, Wang Y, Lv M, Li T, et al. Circular RNA circLMO1 suppresses cervical cancer growth and
metastasis by triggering miR-4291/ACSL4-mediated ferroptosis. Front Oncol. 2022;12:858598. Article CAS PubMed PubMed Central Google Scholar * Liu Z, Wang Q, Wang X, Xu Z, Wei X, Li J.
Circular RNA cIARS regulates ferroptosis in HCC cells through interacting with RNA binding protein ALKBH5. Cell Death Discov. 2020;6:72. Article PubMed PubMed Central Google Scholar *
Jiang L, Kon N, Li T, Wang S, Su T, Hibshoosh H, et al. Ferroptosis as a p53-mediated activity during tumour suppression. Nature. 2015;520:57–62. Article CAS PubMed PubMed Central Google
Scholar * Xie Y, Zhu S, Song X, Xie Y, Zhu S, Song X, et al. The tumor suppressor p53 limits ferroptosis by blocking DPP4 Activity. Cell Rep. 2017;20:1692–704. Article CAS PubMed Google
Scholar * Tarangelo A, Magtanong L, Bieging-rolett KT, Li Y, Ye J, Attardi LD, et al. p53 suppresses metabolic stress-induced ferroptosis in cancer cells. Cell Rep. 2018;22:569–75. Article
CAS PubMed PubMed Central Google Scholar * Gao R, Kalathur RKR, Coto-llerena M, Ercan C, Buechel D, Shuang S, et al. YAP/TAZ and ATF4 drive resistance to Sorafenib in hepatocellular
carcinoma by preventing ferroptosis. EMBO Mol Med. 2021;13:e14351. Article CAS PubMed PubMed Central Google Scholar * Xu Y, Zhang N, Chen C, Xu X, Luo A, Yan Y, et al. Sevoflurane
Induces Ferroptosis of Glioma Cells Through Activating the ATF4-CHAC1 Pathway. Front Oncol. 2022;12:859621. Article CAS PubMed PubMed Central Google Scholar * Wang L, Liu Y, Du T, Yang
H, Lei L, Guo M, et al. ATF3 promotes erastin-induced ferroptosis by suppressing system Xc. Cell Death Differ. 2020;27:662–75. Article CAS PubMed Google Scholar * Harvey KF, Zhang X,
Thomas DM. The Hippo pathway and human cancer. Nat Rev Cancer. 2013;13:246–57. Article CAS PubMed Google Scholar * Nguyen CDK, Yi C. YAP/TAZ Signaling and Resistance to Cancer Therapy.
Trends Cancer. 2019;5:283–96. Article CAS PubMed PubMed Central Google Scholar * Wu J, Minikes AM, Gao M, Bian H, Li Y, Stockwell BR, et al. Intercellular interaction dictates cancer
cell ferroptosis via NF2–YAP signalling. Nature. 2019;572:402–6. Article CAS PubMed PubMed Central Google Scholar * Yang W, Ding CC, Sun T, Rupprecht G, Lin C, Hsu D, et al. The Hippo
pathway effector TAZ regulates ferroptosis in renal cell carcinoma. Cell Rep. 2019;28:2501–2508.e4. Article CAS PubMed Google Scholar * Liu J, Yang M, Kang R, Klionsky DJ, Tang D.
Autophagic degradation of the circadian clock regulator promotes ferroptosis. Autophagy. 2019;15:2033–5. Article CAS PubMed PubMed Central Google Scholar * Yang M, Chen P, Liu J, Zhu S,
Kroemer G. Clockophagy is a novel selective autophagy process favoring ferroptosis. Sci Adv. 2019;5:eaaw2238. Article CAS PubMed PubMed Central Google Scholar * Fan Z, Wirth A, Chen D,
Wruck CJ, Rauh M, Buchfelder M, et al. Nrf2-Keap1 pathway promotes cell proliferation and diminishes ferroptosis. Oncogenesis. 2017;6:e371. Article CAS PubMed PubMed Central Google
Scholar * Carmona-Gutierrez D, Zimmermann A, Kainz K, Pietrocola F, Chen G, Maglioni S, et al. The flavonoid 4,4’-dimethoxychalcone promotes autophagy-dependent longevity across species.
Nat Commun. 2019;10:651. Article PubMed PubMed Central Google Scholar * Yang C, Wang T, Zhao Y, Meng X, Ding W, Wang Q, et al. Flavonoid 4,4'-dimethoxychalcone induced ferroptosis
in cancer cells by synergistically activating Keap1/Nrf2/HMOX1 pathway and inhibiting FECH. Free Radic Biol Med. 2022;188:14–23. Article CAS PubMed Google Scholar * Meng C, Zhan J, Chen
D, Shao G, Zhang H, Gu W, et al. The deubiquitinase USP11 regulates cell proliferation and ferroptotic cell death via stabilization of NRF2 USP11 deubiquitinates and stabilizes NRF2.
Oncogene. 2021;40:1706–20. Article CAS PubMed Google Scholar * Tsai Y, Xia C, Sun Z. The Inhibitory Effect of 6-Gingerol on ubiquitin-specific peptidase 14 enhances autophagy-dependent
ferroptosis and anti-tumor in vivo and in vitro. Front Pharm. 2020;11:598555. Article Google Scholar * Chen S, Bu D, Zhu J, Yue T, Guo S, Wang X, et al. Endogenous hydrogen sulfide
regulates xCT stability through persulfidation of OTUB1 at cysteine 91 in colon cancer cells. Neoplasia. 2021;23:461–72. Article CAS PubMed PubMed Central Google Scholar * Yang Y, Luo
M, Zhang K, Zhang J, Gao T, Connell DO, et al. Nedd4 ubiquitylates VDAC2/3 to suppress erastin-induced ferroptosis in melanoma. Nat Commun. 2020;11:433. Article CAS PubMed PubMed Central
Google Scholar * Zhang HL, Hu BX, Li ZL, Du T, Shan JL, Ye ZP, et al. PKCβII phosphorylates ACSL4 to amplify lipid peroxidation to induce ferroptosis. Nat Cell Biol. 2022;24:88–98.
Article CAS PubMed Google Scholar * Song X, Zhu S, Chen P, Hou W, Wen Q, Liu J, et al. AMPK-Mediated BECN1 Phosphorylation Promotes Ferroptosis by Directly Blocking System Xc– Activity.
Curr Biol. 2018;28:2388–2399.e5. Article CAS PubMed PubMed Central Google Scholar * Zhang X, Sui S, Wang L, Li H, Zhang L, Xu S, et al. Inhibition of tumor propellant glutathione
peroxidase 4 induces ferroptosis in cancer cells and enhances anticancer effect of cisplatin. J Cell Physiol. 2020;235:3425–37. Article CAS PubMed Google Scholar * Logie E, Van Puyvelde
B, Cuypers B, Schepers A, Berghmans H, Verdonck J, et al. Ferroptosis induction in multiple myeloma cells triggers DNA methylation and histone modification changes associated with cellular
senescence. Int J Mol Sci. 2021;22:12234. Article CAS PubMed PubMed Central Google Scholar * Hasegawa M, Takahashi H, Rajabi H, Alam M, Suzuki Y, Yin L, et al. Functional interactions
of the cystine/glutamate antiporter, CD44V and MUC1-C oncoprotein in triple-negative breast cancer cells. Oncotarget. 2016;7:11756–69. Article PubMed PubMed Central Google Scholar * Wang
S, Li D, Ou Y, Wang S, Li D, Ou Y, et al. Acetylation is crucial for p53-mediated ferroptosis and tumor suppression. Cell Rep. 2016;17:366–73. Article CAS PubMed PubMed Central Google
Scholar * Birsen R, Larrue C, Decroocq J, Johnson N, Guiraud N, Gotanegre M, et al. APR-246 induces early cell death by ferroptosis in acute myeloid leukemia. Haematologica.
2022;107:403–16. Article CAS PubMed Google Scholar * Jin L, Tong L. PAQR3 inhibits proliferation and aggravates ferroptosis in acute lymphoblastic leukemia through modulation Nrf2
stability. Immun Inflamm Dis. 2021;9:827–39. Article CAS PubMed PubMed Central Google Scholar * Tang Z, Jiang W, Mao M, Zhao J, Chen J, Cheng N. Deubiquitinase USP35 modulates
ferroptosis in lung cancer via targeting ferroportin. Clin Transl Med. 2021;11:e390. Article CAS PubMed PubMed Central Google Scholar * Yang J, Zhou Y, Xie S, Wang J, Li Z, Chen L, et
al. Metformin induces Ferroptosis by inhibiting UFMylation of SLC7A11 in breast cancer. J Exp Clin Cancer Res. 2021;40:206. Article CAS PubMed PubMed Central Google Scholar * Ye Z, Zhuo
Q, Hu Q, Xu X, Liu M, Zhang Z, et al. FBW7-NRA41-SCD1 axis synchronously regulates apoptosis and ferroptosis in pancreatic cancer cells. Redox Biol. 2021;38:101807. Article CAS PubMed
Google Scholar * Yang Y, Lin J, Guo S, Xue X, Wang Y, Qiu S, et al. RRM2 protects against ferroptosis and is a tumor biomarker for liver cancer. Cancer Cell Int. 2020;20:587. Article
PubMed PubMed Central Google Scholar * Zhu G, Murshed A, Li H, Ma J, Zhen N, Ding M, et al. O-GlcNAcylation enhances sensitivity to RSL3-induced ferroptosis via the YAP/TFRC pathway in
liver cancer. Cell Death Discov. 2021;7:83. Article CAS PubMed PubMed Central Google Scholar * Jiang B, Zhao YQ, Shi M, Song L, Wang Q, Qin QM, et al. DNAJB6 promotes ferroptosis in
esophageal squamous cell carcinoma. Dig Dis Sci. 2020;65:1999–2008. Article CAS PubMed Google Scholar * Wang J, Wang T, Zhang Y, Liu J, Song J. CPEB1 enhances erastin-induced ferroptosis
in gastric cancer cells by suppressing twist1 expression. IUBMB Life. 2021;73:1180–90. Article CAS PubMed Google Scholar * Yao F, Cui X, Zhang Y, Bei Z, Wang H, Zhao D, et al. Iron
regulatory protein 1 promotes ferroptosis by sustaining cellular iron homeostasis in melanoma. Oncol Lett. 2021;22:657. Article CAS PubMed PubMed Central Google Scholar * Son J,
Lyssiotis CA, Ying H, Wang X, Hua S, Ligorio M, et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature. 2013;496:101–5. Article CAS PubMed
PubMed Central Google Scholar * Kremer DM, Nelson BS, Lin L, Yarosz EL, Halbrook CJ, Kerk SA, et al. GOT1 inhibition promotes pancreatic cancer cell death by ferroptosis. Nat Commun.
2021;12:4860. Article CAS PubMed PubMed Central Google Scholar * Lu Y, Qin H, Jiang B, Lu W, Hao J, Cao W, et al. KLF2 inhibits cancer cell migration and invasion by regulating
ferroptosis through GPX4 in clear cell renal cell carcinoma. Cancer Lett. 2021;522:1–13. Article CAS PubMed Google Scholar * Xu F, Guan Y, Gao M, Chong T. The roles of ferroptosis
regulatory gene SLC7A11 in renal cell carcinoma: a multi-omics study. Cancer Med. 2021;10:9078–96. Article CAS PubMed PubMed Central Google Scholar * Lu D, Yang Z, Xia Q, Gao S, Sun S,
Luo X, et al. ACADSB regulates ferroptosis and affects the migration, invasion and proliferation of colorectal cancer cells. Cell Biol Int. 2020;44:2334–43. Article CAS PubMed Google
Scholar * Qin X, Zhang J, Lin Y, Sun XM, Zhang JN, Cheng ZQ. Identification of MiR-211-5p as a tumor suppressor by targeting ACSL4 in hepatocellular carcinoma. J Transl Med. 2020;18:326.
Article CAS PubMed PubMed Central Google Scholar * Feng L, Zhao K, Sun L, Yin X, Zhang J, Liu C, et al. SLC7A11 regulated by NRF2 modulates esophageal squamous cell carcinoma
radiosensitivity by inhibiting ferroptosis. J Transl Med. 2021;19:367. Article CAS PubMed PubMed Central Google Scholar * He Q, Liu M, Huang W, Chen X, Zhang B, Zhang T, et al.
IL-1β-induced elevation of solute carrier family 7 member 11 promotes hepatocellular carcinoma metastasis through up-regulating programmed death ligand 1 and colony-stimulating factor 1.
Hepatology. 2021;74:3174–93. Article CAS PubMed Google Scholar * Liu W, Chakraborty B, Safi R, Kazmin D, Chang CY, McDonnell DP. Dysregulated cholesterol homeostasis results in
resistance to ferroptosis increasing tumorigenicity and metastasis in cancer. Nat Commun. 2021;12:5103. Article CAS PubMed PubMed Central Google Scholar * Nagpal A, Redvers RP, Ling X,
Ayton S, Fuentes M, Tavancheh E, et al. Neoadjuvant neratinib promotes ferroptosis and inhibits brain metastasis in a novel syngeneic model of spontaneous HER2+ve breast cancer metastasis.
Breast Cancer Res. 2019;21:94. Article PubMed PubMed Central Google Scholar * Jansen RS, Mahakena S, de Haas M, Borst P, van de Wetering K. ATP-binding cassette subfamily c member 5
(ABCC5) functions as an Efflux transporter of glutamate conjugates and analogs. J Biol Chem. 2015;290:30429–40. Article CAS PubMed PubMed Central Google Scholar * Huang W, Chen K, Lu Y,
Zhang D, Cheng Y, Li L, et al. ABCC5 facilitates the acquired resistance of sorafenib through the inhibition of SLC7A11-induced ferroptosis in hepatocellular carcinoma. Neoplasia.
2021;23:1227–39. Article CAS PubMed PubMed Central Google Scholar * Rusolo F, Capone F, Pasquale R, Angiolillo A, Colonna G, Castello G, et al. Comparison of the seleno-transcriptome
expression between human non-cancerous mammary epithelial cells and two human breast cancer cell lines. Oncol Lett. 2017;13:2411–7. Article CAS PubMed PubMed Central Google Scholar *
Dinarvand N, Khanahmad H, Hakimian SM. Evaluation of long-chain acyl-coenzyme A synthetase 4 (ACSL4) expression in human breast cancer. Res Pharm Sci. 2020;15:48–56. Article PubMed PubMed
Central Google Scholar * Sha R, Xu Y, Yuan C, Sheng X, Wu Z, Peng J, et al. Predictive and prognostic impact of ferroptosis-related genes ACSL4 and GPX4 on breast cancer treated with
neoadjuvant chemotherapy. EBioMedicine. 2021;71:103560. Article CAS PubMed PubMed Central Google Scholar * Dai E, Han L, Liu J, Xie Y, Zeh HJ, Kang R, et al. Ferroptotic damage promotes
pancreatic tumorigenesis through a TMEM173/STING-dependent DNA sensor pathway. Nat Commun. 2020;11:6339. Article CAS PubMed PubMed Central Google Scholar * Ma X, Xiao L, Liu L, Ye L,
Su P, Bi E, et al. CD36-mediated ferroptosis dampens intratumoral CD8+ T cell effector function and impairs their antitumor ability. Cell Metab. 2021;33:1001–1012.e5. Article CAS PubMed
PubMed Central Google Scholar * Lachaier E, Louandre C, Godin C, Saidak Z, Baert M, Diouf M, et al. Sorafenib induces ferroptosis in human cancer cell lines originating from different
solid tumors. Anticancer Res. 2014;34:6417–22. CAS PubMed Google Scholar * Wang Q, Bin C, Xue Q, Gao Q, Huang A, Wang K, et al. GSTZ1 sensitizes hepatocellular carcinoma cells to
sorafenib-induced ferroptosis via inhibition of NRF2/GPX4 axis. Cell Death Dis. 2021;12:426. Article PubMed PubMed Central Google Scholar * Ghosh S. Cisplatin: The first metal based
anticancer drug. Bioorg Chem. 2019;88:102925. Article CAS PubMed Google Scholar * Ghoochani A, Hsu EC, Aslan M, Rice MA, Nguyen HM, Brooks JD, et al. Ferroptosis inducers are a novel
therapeutic approach for advanced prostate cancer. Cancer Res. 2021;81:1583–94. Article CAS PubMed PubMed Central Google Scholar * Wang J, Liu Z, Wang Z, Wang S, Chen Z, Li Z, et al.
Targeting c-Myc: JQ1 as a promising option for c-Myc-amplified esophageal squamous cell carcinoma. Cancer Lett. 2018;419:64–74. Article CAS PubMed Google Scholar * Jiang G, Deng W, Liu
Y, Wang C. General mechanism of JQ1 in inhibiting various types of cancer. Mol Med Rep. 2020;21:1021–34. CAS PubMed PubMed Central Google Scholar * Sui S, Zhang J, Xu S, Wang Q, Wang P,
Pang D. Ferritinophagy is required for the induction of ferroptosis by the bromodomain protein BRD4 inhibitor (+)-JQ1 in cancer cells. Cell Death Dis. 2019;10:331. Article PubMed PubMed
Central Google Scholar * Yang L, Chen X, Yang Q, Chen J, Huang Q, Yao L, et al. Broad spectrum deubiquitinase inhibition induces both apoptosis and ferroptosis in cancer cells. Front
Oncol. 2020;10:949. Article PubMed PubMed Central Google Scholar * Sun D, Li YC, Zhang XY. Lidocaine promoted ferroptosis by targeting miR-382-5p /SLC7A11 axis in ovarian and breast
cancer. Front Pharm. 2021;12:681223. Article CAS Google Scholar * Hao J, Zhang W, Huang Z. Bupivacaine modulates the apoptosis and ferroptosis in bladder cancer via phosphatidylinositol
3-kinase (PI3K)/AKT pathway. Bioengineered. 2022;13:6794–806. Article CAS PubMed PubMed Central Google Scholar * Fallon MT, Wilcock A, Kelly CA, Paul J, Lewsley LA, Norrie J, et al.
Oral ketamine vs placebo in patients with cancer-related neuropathic pain: A randomized clinical trial. JAMA Oncol. 2018;4:870–2. Article PubMed PubMed Central Google Scholar * Li J,
Lama R, Galster SL, Inigo JR, Wu J, Chemler SR, et al. Small molecule MMRi62 induces ferroptosis and inhibits metastasis in pancreatic cancer via degradation of ferritin heavy chain and
mutant p53. Mol Cancer Ther. 2022;21:535–45. Article CAS PubMed PubMed Central Google Scholar * Shanmugam MK, Warrier S, Kumar AP, Sethi G, Arfuso F. Potential role of natural compounds
as anti-angiogenic agents in cancer. Curr Vasc Pharm. 2017;15:503–19. Article CAS Google Scholar * Koch W, Kukula-Koch W, Marzec Z, Kasperek E, Wyszogrodzka-Koma L, Szwerc W, et al.
Application of chromatographic and spectroscopic methods towards the quality assessment of ginger (Zingiber officinale) rhizomes from ecological plantations. Int J Mol Sci. 2017;18:452.
Article PubMed PubMed Central Google Scholar * Zhang F, Ma N, Gao YF, Sun LL, Zhang JG. Therapeutic effects of 6-Gingerol, 8-Gingerol, and 10-Gingerol on dextran sulfate sodium-induced
acute ulcerative colitis in rats. Phyther Res. 2017;31:1427–32. Article CAS Google Scholar * de Lima RMT, Dos Reis AC, de Menezes APM, Santos JVO, Filho JWGO, Ferreira JRO, et al.
Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phyther Res. 2018;32:1885–907. Article Google Scholar *
Ding Y, Chen X, Liu C, Ge W, Wang Q, Hao X, et al. Identification of a small molecule as inducer of ferroptosis and apoptosis through ubiquitination of GPX4 in triple negative breast cancer
cells. J Hematol Oncol. 2021;14:19. Article CAS PubMed PubMed Central Google Scholar * Zhang W, Jiang B, Liu Y, Xu L, Wan M. Bufotalin induces ferroptosis in non-small cell lung cancer
cells by facilitating the ubiquitination and degradation of GPX4. Free Radic Biol Med. 2022;180:75–84. Article CAS PubMed Google Scholar * Bai Y, Yang L, Zhang C, Yang Y. Studies on the
Mechanism of alloimperatorin on the proliferation and apoptosis of hela cells. J Oncol. 2021;2021:6617312. Article PubMed PubMed Central Google Scholar * Zhang J, Gao R, Li J, Yu K, Bi
K. Alloimperatorin activates apoptosis, ferroptosis and oxeiptosis to inhibit the growth and invasion of breast cancer cells in vitro. Biochem Cell Biol. 2022;100:213–22. Article CAS
PubMed Google Scholar * Lang X, Green MD, Wang W, Yu J, Choi JE, Jiang L, et al. Radiotherapy and immunotherapy promote tumoral lipid oxidation and ferroptosis via synergistic repression
of SLC7A11. Cancer Discov. 2019;9:1673–85. Article CAS PubMed PubMed Central Google Scholar * Ye LF, Chaudhary KR, Zandkarimi F, Harken AD, Kinslow CJ, Upadhyayula S, et al.
Radiation-induced lipid peroxidation triggers ferroptosis and synergizes with ferroptosis inducers. ACS Chem Biol. 2020;15:469–84. Article CAS PubMed PubMed Central Google Scholar * Lei
G, Zhang Y, Koppula P, Liu X, Zhang J, Lin SH, et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression. Cell Res. 2020;30:146–62. Article CAS PubMed
PubMed Central Google Scholar * Wang S, Chen S, Ying Y, Ma X, Shen H, Li J, et al. Comprehensive analysis of ferroptosis regulators with regard to PD-L1 and immune infiltration in clear
cell renal cell carcinoma. Front Cell Dev Biol. 2021;9:676142. Article PubMed PubMed Central Google Scholar * Liao P, Wang W, Wang W, Kryczek I, Li X, Bian Y, et al. CD8+ T cells and
fatty acids orchestrate tumor ferroptosis and immunity via ACSL4. Cancer Cell. 2022;40:365–378.e6. Article CAS PubMed PubMed Central Google Scholar * Song R, Li T, Ye J, Sun F, Hou B,
Saeed M, et al. Acidity-activatable dynamic nanoparticles boosting ferroptotic cell death for immunotherapy of cancer. Adv Mater. 2021;33:e2101155. Article PubMed Google Scholar * Wang W,
Green M, Choi JE, Gijón M, Kennedy PD, Johnson JK, et al. CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. Nature. 2019;569:270–4. Article CAS PubMed PubMed Central
Google Scholar * Matsushita M, Freigang S, Schneider C, Conrad M, Bornkamm GW, Kopf M. T cell lipid peroxidation induces ferroptosis and prevents immunity to infection. J Exp Med.
2015;212:555–68. Article CAS PubMed PubMed Central Google Scholar * Yang Y, Wang Y, Guo L, Gao W, Tang T, Yan M. Interaction between macrophages and ferroptosis. Cell Death Dis.
2022;13:355. Article CAS PubMed PubMed Central Google Scholar * Liu Z, Ma Y, Cui Q, Xu J, Tang Z, Wang Y, et al. Toll-like receptor 4 plays a key role in advanced glycation end
products-induced M1 macrophage polarization. Biochem Biophys Res Commun. 2020;531:602–8. Article CAS PubMed Google Scholar * Zhou Y, Que KT, Zhang Z, Yi ZJ, Zhao PX, You Y, et al. Iron
overloaded polarizes macrophage to proinflammation phenotype through ROS/acetyl-p53 pathway. Cancer Med. 2018;7:4012–22. Article CAS PubMed PubMed Central Google Scholar * Hu X, Cai X,
Ma R, Fu W, Zhang C, Du X. Iron-load exacerbates the severity of atherosclerosis via inducing inflammation and enhancing the glycolysis in macrophages. J Cell Physiol. 2019;234:18792–18800.
Article CAS PubMed Google Scholar * Hao X, Zheng Z, Liu H, Zhang Y, Kang J, Kong X, et al. Inhibition of APOC1 promotes the transformation of M2 into M1 macrophages via the ferroptosis
pathway and enhances anti -PD1 immunotherapy in hepatocellular carcinoma based on single-cell RNA sequencing. Redox Biol. 2022;56:102463. Article CAS PubMed PubMed Central Google Scholar
* Tang W, Xu F, Zhao M, Zhang S. Ferroptosis regulators, especially SQLE, play an important role in prognosis, progression and immune environment of breast cancer. BMC Cancer.
2021;21:1160. Article CAS PubMed PubMed Central Google Scholar * Wu L, Yin L, Gu Y. The establishment of polypeptide PSMA-targeted chimeric antigen receptor-engineered natural killer
cells for castration-resistant prostate cancer and the induction of ferroptosis-related cell death. Cancer Commun (Lond). 2022;42:768–83. Article PubMed Google Scholar * Kim SE, Zhang L,
Ma K, Riegman M, Chen F, Ingold I, et al. Ultrasmall nanoparticles induce ferroptosis in nutrient-deprived cancer cells and suppress tumour growth. Nat Nanotechnol. 2016;11:977–85. Article
CAS PubMed PubMed Central Google Scholar * Gabizon A, Bradbury M, Prabhakar U, Zamboni W, Libutti S, Grodzinski P. Cancer nanomedicines: closing the translational gap. Lancet.
2014;384:2175–6. Article PubMed PubMed Central Google Scholar * Zhang Y, Tan H, Daniels JD, Uchida K, Connor OAO, Stockwell BR, et al. Imidazole ketone erastin induces ferroptosis and
slows tumor growth in a mouse lymphoma model. Cell Chem Biol. 2019;26:623–33. Article CAS PubMed PubMed Central Google Scholar * Li Z, Wang C, Dai C, Hu R, Ding L, Feng W, et al.
Biomaterials engineering dual catalytic nanomedicine for autophagy-augmented and ferroptosis-involved cancer nanotherapy. Biomaterials. 2022;287:121668. Article CAS PubMed Google Scholar
* Wei F, Zhong L, Li X, Meng D, Hui H, Cai D, et al. A Sonication-activated valence-variable sono-sensitizer/catalyst for autography inhibition/ ferroptosis-induced tumor nanotherapy.
Angew Chem Int Ed Engl. 2022;61:e202212021. Google Scholar * Hu Z, Song X, Ding L, Cai Y, Yu L, Zhang L, et al. Engineering Fe/Mn-doped zinc oxide nanosonosensitizers for
ultrasound-activated and multiple ferroptosis-augmented nanodynamic tumor suppression. Mater Today Bio. 2022;16:100452. Article CAS PubMed PubMed Central Google Scholar * Zhu M, Wu P,
Li Y, Zhang L, Zong Y, Wan M. Synergistic therapy for orthotopic gliomas via biomimetic nanosonosensitizer-mediated sonodynamic therapy and ferroptosis. Biomater Sci. 2022;10:3911–23.
Article CAS PubMed Google Scholar * Edition I. Self-assembled Aza-Boron-Dipyrromethene for ferroptosis-boosted sonodynamic therapy. Angew Chem. 2022;61:e202210174. Google Scholar * Xu
T, Ma Y, Yuan Q, Hu H, Hu X, Qian Z, et al. Enhanced ferroptosis by oxygen-boosted phototherapy based on 2- in-1 nanoplatform of ferrous hemoglobin for tumor synergistic therapy.
Theranostics. 2019;18:3293–307. Google Scholar * Li L, Song D, Qi L, Jiang M, Wu Y, Gan J, et al. Photodynamic therapy induces human esophageal carcinoma cell pyroptosis by targeting the
PKM2 / caspase-8 / caspase-3 / GSDME axis. Cancer Lett. 2021;520:143–59. Article CAS PubMed Google Scholar * Moghissi K, Hons KDBA. Update on the current indications, practice and
resultsof photodynamic therapy (PDT) in early central lung cancer (ECLC.Photodiagnosis Photodyn Ther. 2008;5:10–18. * Hamdoon Z, Jerjes W, Rashed D, Kawas S, Samsudin R, Hopper C.
Photodiagnosis and photodynamic therapy in vivo optical coherence tomography-guided photodynamic therapy for skin pre-cancer and cancer. Photodiagnosis Photodyn Ther. 2021;12:102520. Article
Google Scholar * Wu M, Ling W, Wei J, Liao R, Sun H, Li D. Biomimetic photosensitizer nanocrystals trigger enhanced ferroptosis for improving cancer treatment. J Control Release.
2022;352:1116–33. Article CAS PubMed Google Scholar * Liang X, Chen M, Bhattarai P, Hameed S, Tang Y, Dai Z. Complementing cancer photodynamic therapy with ferroptosis through iron oxide
loaded porphyrin-grafted lipid nanoparticles. ACS Nano. 2021;15:20164–80. Article CAS PubMed Google Scholar * Ouyang S, Li H, Lou L, Huang Q, Zhang Z, Mo J, et al. Inhibition of
STAT3-ferroptosis negative regulatory axis suppresses tumor growth and alleviates chemoresistance in gastric cancer. Redox Biol. 2022;52:102317. Article CAS PubMed PubMed Central Google
Scholar * Badgley MA, Kremer DM, Maurer HC, Delgiorno KE, Lee H, Purohit V, et al. Cysteine depletion induces pancreatic tumor ferroptosis in mice. Science 2020;89:85–89. Article Google
Scholar * Sun J, Zhou C, Zhao Y, Zhang X, Chen W, Zhou Q, et al. Quiescin sulfhydryl oxidase 1 promotes sorafenib-induced ferroptosis in hepatocellular carcinoma by driving EGFR endosomal
trafficking and inhibiting NRF2 activation. Redox Biol. 2021;41:101942. Article CAS PubMed PubMed Central Google Scholar Download references ACKNOWLEDGEMENTS Figure 1, Fig. 2 and Fig. 3
were created under the academic license of BioRender.com (Invoice number: GE25C8PVXY, JD25C8PX01, RN25C8PUKH). FUNDING The work was supported by the National Natural Science Foundation of
China (81972330 to X.L., 82002731 to T.H.); Major science and technology project of Xinxiang City (21ZD013 to X.L.); the Henan National Science Fund for Excellent Young Scholars
(212300410067 to T.H.). AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Gastroenterology, the Third Affiliated Hospital of Xinxiang Medical University, Henan Key Laboratory of
Tumor Molecular Therapy Medicine, Xinxiang, 453003, Henan Province, China Zhen Chen, Weilong Wang, Tao Han & Xiumin Li * Department of Biomedical Science, Advanced Medical and Dental
Institute, Universiti Sains Malaysia, Bertam 13200, Kepala Batas, Pulau Pinang, Malaysia Zhen Chen, Weilong Wang, Siti Razila Abdul Razak & Nor Hazwani Ahmad * Xinxiang Key Laboratory
for Molecular Therapy of Cancer, Xinxiang Medical University, Xinxiang, 453003, Henan Province, P. R. China Zhen Chen, Weilong Wang, Tao Han & Xiumin Li * Institutes of Health Central
Plains, Xinxiang Medical University, Xinxiang, 453003, Henan Province, P. R. China Tao Han Authors * Zhen Chen View author publications You can also search for this author inPubMed Google
Scholar * Weilong Wang View author publications You can also search for this author inPubMed Google Scholar * Siti Razila Abdul Razak View author publications You can also search for this
author inPubMed Google Scholar * Tao Han View author publications You can also search for this author inPubMed Google Scholar * Nor Hazwani Ahmad View author publications You can also search
for this author inPubMed Google Scholar * Xiumin Li View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS ZC and WW drafted the original
manuscript. ZC designed and created the figures and tables. NHA and XL designed and revised the manuscript. TH and SRAR participated in the revision of the manuscript. CORRESPONDING AUTHORS
Correspondence to Nor Hazwani Ahmad or Xiumin Li. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER’S NOTE Springer Nature
remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. Edited by Boris Zhivotovsky SUPPLEMENTARY INFORMATION AJ-CHECKLIST RIGHTS AND
PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any
medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The
images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not
included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly
from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Chen, Z., Wang,
W., Abdul Razak, S.R. _et al._ Ferroptosis as a potential target for cancer therapy. _Cell Death Dis_ 14, 460 (2023). https://doi.org/10.1038/s41419-023-05930-w Download citation * Received:
24 January 2023 * Revised: 24 May 2023 * Accepted: 23 June 2023 * Published: 24 July 2023 * DOI: https://doi.org/10.1038/s41419-023-05930-w SHARE THIS ARTICLE Anyone you share the following
link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature
SharedIt content-sharing initiative