Play all audios:
ABSTRACT The history of the Earth has been marked by major ecological transitions, driven by metabolic innovation, that radically reshaped the composition of the oceans and atmosphere. The
nature and magnitude of the earliest transitions, hundreds of million years before photosynthesis evolved, remain poorly understood. Using a novel ecosystem-planetary model, we find that
pre-photosynthetic methane-cycling microbial ecosystems are much less productive than previously thought. In spite of their low productivity, the evolution of methanogenic metabolisms
strongly modifies the atmospheric composition, leading to a warmer but less resilient climate. As the abiotic carbon cycle responds, further metabolic evolution (anaerobic methanotrophy) may
feed back to the atmosphere and destabilize the climate, triggering a transient global glaciation. Although early metabolic evolution may cause strong climatic instability, a low CO:CH4
atmospheric ratio emerges as a robust signature of simple methane-cycling ecosystems on a globally reduced planet such as the late Hadean/early Archean Earth. SIMILAR CONTENT BEING VIEWED BY
OTHERS CARBON ISOTOPE EVIDENCE FOR LARGE METHANE EMISSIONS TO THE PROTEROZOIC ATMOSPHERE Article Open access 23 October 2020 METHANE FORMATION DRIVEN BY LIGHT AND HEAT PRIOR TO THE ORIGIN
OF LIFE AND BEYOND Article Open access 01 August 2023 THE MICROBIAL CARBON PUMP AND CLIMATE CHANGE Article 15 March 2024 INTRODUCTION By 3.5 Ga, life had emerged on Earth1,2,3. Astrophysical
and geophysical data concur in showing that the planet was habitable 400 My earlier at the very least, and possibly as early as ~4.5 Gya, depending on the occurrence, magnitude, and effect
of large asteroid impacts during the Hadean3. Early on, Earth’s carbon cycle likely established and maintained temperate climatic conditions4,5 in spite of a Sun being 20–25% dimmer than it
is today6. The earliest microbial ecosystems evolving under these conditions, hundreds of million years before the first anoxygenic phototrophs7 became actors of the Archean climate8, most
likely involved chemolithotrophs (i.e., unicellular organisms that use redox potential as energy source for biomass production) producing methane as a metabolic waste. Phylogenetic
analyses2,7,9,10 combined with isotopic evidence11 and the then-time predominance12,13 of the electron donors H2 and CO lend weight to a very early origin of H2-based methanogens (MG),
CO-based autotrophic acetogens (AG), and methanogenic acetotrophs (AT). As CH4 built up in the atmosphere, the evolution of anaerobic methanotrophy (MT) may have been favored. Contrary to
the modern biosphere, the biomass productivity of such ecosystems was likely low and energy limited, i.e., limited by the availability of electron donors rather than nutrients such as
nitrogen, phosphorus, or iron13,14. Whether and how the evolution of a primitive biosphere formed by these metabolisms influenced the planetary environment globally is unclear. Previous
studies8,12,13 have addressed the productivity of primitive, chemolithotrophic ecosystems and their influence on the young Earth’s equilibrium atmospheric conditions. Such studies relied on
equilibrium analyses of the planetary ecosystem; they made strongly simplifying assumptions on the function of chemolithotrophic microbial metabolisms, and did not close the feedback loop
linking biological activity, atmospheric composition, and climate. Although these studies showed that primitive biospheres may have had a significant impact on the planet’s early atmosphere
and climate, their ability to quantify this impact and estimate the underlying biomass productivity was limited. Furthermore, earlier theory based on equilibrium analyses could not address
the coupled dynamics of metabolic evolution and planetary surface conditions, whereby evolutionary changes might trigger significant atmospheric and climatic events and lead to novel steady
states. Thus, advancing existing models is needed to generate hypotheses on the history of atmospheric and climatic conditions that metabolic evolutionary innovation may have driven on the
early Earth. Here we ask, how constrained was the habitability of late Hadean/early Archean Earth to methane-cycling ecosystems? How productive were these ecosystems and did they have a
significant impact on the atmospheric composition and climate? How did their impact change as different metabolisms evolved? To answer these questions, we lay out a new probabilistic
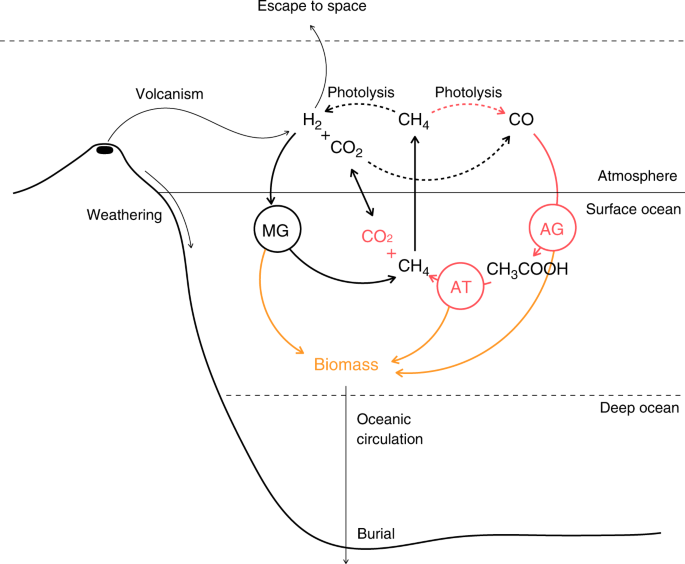
modeling framework for an evolving microbial community coupled to early Earth surface geochemistry and climate (Fig. 1, see “Methods” and Supplementary “Results and Discussion” for further
details). The mean surface temperature and the composition of the atmosphere and oceans are parameterized using a 3D climate model4,15 and a 1D photochemical model16, combined with a simple
temperature-dependent carbon cycle model5. Advancing the previous studies8,12,13, this planetary model is coupled dynamically to a biological model of cell population dynamics and
evolutionary adaptation, constructed by scaling the intracellular processes of energy acquisition (i.e., catabolism), cell maintenance, and biomass production (i.e., anabolism) up to
ecosystem function17,18. The biological model is grounded in thermodynamics and based on observations of cell size and temperature kinetic dependencies, widely and robustly shared among
modern unicellular organisms19,20,21,22,23. Quantitative validation of similar models was obtained from laboratory experiments on anoxic ecosystems in bioreactors18. RESULTS ECOSYSTEMS
VIABILITY First, we assess the viability of the methanogenic biospheres (MG, AG + AT, or MG + AG + AT) on an initially cool, lifeless Earth. The initial abiotic surface temperature _T_Geo is
assumed to be 12 °C, corresponding to _p_CO2 = 2 × 105 ppm, negligible _p_CH4, and volcanic outgassing of H2, _φ_volc(H2), ranging from 5 × 109 to 2 × 1011 molecules cm−2 s−1. By performing
a Monte-Carlo exploration of the space of biological parameters, we generate posterior distributions of possible life-atmosphere-climate outcomes (Supplementary Figs. 1 and 2), thus
providing general insights that do not depend on specific parameterizations of chemolithotrophic metabolisms. We find that all three methanogenic ecosystems are viable (i.e., they can
sustain a steady, positive biomass production) in more than 50% of the simulations, regardless of the intensity of H2 outgassing (further information on the region of viability in the space
of biological parameters is provided in the Supplementary Results and Discussion). Among viable ecosystems, the posterior distributions of the planetary and ecological state variables are
peaked and relatively narrow (Fig. 2). Hereafter we will focus on median values to describe model outputs. SHORT-TERM EFFECTS OF METABOLIC EVOLUTION We first consider the direct effects on
the early Earth’s atmosphere and climate, on a relatively short timescale of ~106 years, of the transition from the initially cool, lifeless state to a planet populated by one of three
methanogenic biospheres, MG, AG + AT, or MG + AG + AT. On such a short timescale, the carbon cycle has a negligible influence on the atmospheric CO2. H2 is both a metabolic substrate of MG
and involved in the photochemical production of CO, the metabolic substrate of AG (Fig. 1). As a consequence, stronger H2 volcanic outgassing always enhances biomass production and CH4
emission (Fig. 2). The highest CH4 emission is achieved by MG ecosystems, with _p_CH4 ranging from 80 to 4000 ppm and equilibrium temperature, and _T_BioGeo, raised by +7° to +17° (Fig. 2).
The environmental impact of AG + AT ecosystems is similar in magnitude, with _p_CH4 ranging from 50 to 1000 ppm, and temperature increases of +6° to +15° (Fig. 2). The planet’s abiotic
surface temperature, _T_Geo, is likely to have a strong influence on methanogenic activity (see “Methods”). Temperature influences both cell kinetics (metabolisms are slower at lower
temperature) and thermodynamics (strong negative effect of high temperatures on MG, less so on AG + AT). _T_Geo also correlates positively with _p_CO2 and _p_CO, which are both substrates of
methanogenesis. We evaluate the influence of _T_Geo by examining bio-geo environmental feedbacks (i.e., how the change in metabolic activity due to variation in _T_Geo feeds back to
climate) for _T_Geo ranging from −18 to 57 °C, which corresponds to _p_CO2 ranging from 5 × 10-4 to 1 bar (Fig. 3). We ran simulations using a default biological parameterization for which
CH4 emissions are close to the median predictions described above. Note that we also considered _T_Geo as varying independently of _p_CO2, due to e.g. variation in stellar radiation;
corresponding results are shown in Supplementary Fig. 3. If the overall effect of _T_Geo on methanogenic activity is positive, the methanogenic ecosystem is expected to amplify temperature
fluctuations driven by external events such as variation in CO2 outgassing. In this case, increasing _T_Geo should enhance the biogenic emission of CH4, further warming the planet through
additional greenhouse effect; decreasing _T_Geo should have the opposite effect. In contrast, if the overall effect of _T_Geo on methanogenic activity is negative, methanogenic ecosystems
will buffer temperature variation. For the MG ecosystem (Fig. 3), we find that there is a critical abiotic temperature _T_Geo ≈ 5 °C, almost insensitive to H2 outgassing, at which the
warming effect of the ecosystem is maximum, from +5° to +17° across the range of H2 outgassing rates. The MG ecosystem buffers fluctuations of abiotic temperature above the critical value,
whereas it amplifies abiotic temperature fluctuations below the critical value. In contrast, the AG + AT ecosystem always amplifies temperature variation at all abiotic temperatures, and its
influence tends to dominate when MG and AG + AT metabolisms co-occur (Fig. 3). Such amplification or buffering of temperature variations can represent up to 33% of the abiotic fluctuations
(Supplementary Fig. 4). Overall, the function and evolution of methanogenic ecosystems lead to a less resilient, more variable climate. The formation of organic hazes when the
_p_CH4-to-_p_CO2 ratio is greater than 0.2 (ref. 24) has been proposed as a general mechanism of climate regulation25. In the late Archean, _p_CO2 was low5, favoring organic haze formation
that may have prevented hot runaway scenarios. In the Hadean/early Archean, however, our model predicts organic hazes to form in a limited range of conditions, at very high H2 volcanic
outgassing rates, low _p_CO2, and low abiotic temperature close to or below the freezing point (Fig. 3). Under these specific conditions, the formation of organic hazes may overwhelm the
warming effect of methanogenic ecosystems and leave the planet in a globally glaciated state. Under most conditions, however, it is the availability of electron donors (H2, CO) to
methanogenesis, and not organic hazes, that is expected to limit Archean climate warming by biological activity. BIOMASS PRODUCTION In spite of their strong impact on the planet’s atmosphere
and climate, primitive methanogenic ecosystems are characterized by extremely low biomass productivity, AG + AT being the most productive pathway by far (Fig. 2). As microbial
chemolithotrophs consume atmospheric electron donors, they drive the system closer to its thermodynamic equilibrium, thus gradually decreasing the thermodynamic efficiency of the metabolic
coupling between energy acquisition (catabolism) and biomass production (anabolism; see “Methods equation (E2)”). As a consequence, for the highest value of abiotic H2 outgassing, our model
predicts biomass production to range from 106 to 109 molecule C cm−2 s−1. This is 1–4 orders of magnitude below previous estimates13 (based on models that assumed a fixed biomass yield per
electron donor consumed) and 4–7 orders of magnitude below modern values26. Albeit extremely low, biomass production is very sensitive to the metabolic composition of the ecosystem and
temperature, _T_Geo. Supplementary Fig. 5 shows how biomass production is influenced by these two factors. For the MG ecosystem, biomass production peaks for _T_Geo between −10 and 10 °C
depending on the intensity of H2 volcanic outgassing (slightly above 109 molecules C cm−2 s−1 at high rates of H2 volcanic outgassing), and strongly decreases for higher _T_Geo. The maximum
biomass production is of the same order in the AG + AT ecosystem and reached for similar conditions (intermediate _T_Geo, high H2 volcanic outgassing rate), but its dependence upon _T_Geo
and the rate of H2 volcanic outgassing is much weaker. In the MG + AG + AT ecosystem the two methanogenic pathways interact synergistically, leading to a nonlinear, multiplicative increase
in biomass production at low and high temperature. The synergy involves the combination of biogeochemical recycling loops, both locally and globally. Locally, while MG consumes CO2 to
produce CH4, AT decomposes CH3COOH and produces CO2. The metabolic waste of AT is, therefore, the metabolic substrate of MG, and the combination of the two metabolic pathways pulls the
system further away from its thermodynamic equilibrium, hence an increase in the efficiency of both pathways. Globally, as the MG metabolism releases additional CH4 in the atmosphere, the
production of CO through photochemistry is accelerated; CO being the metabolic substrate of AG, its metabolic efficiency is enhanced. The synergistic effect is greatest at low abiotic
temperature. In those very specific conditions and for the highest values of H2 outgassing, biomass production can reach about 1010 molecules C cm−2 s−1—about 1000 times less than estimates
of modern primary production26. METABOLIC EVOLUTION AND THE CARBON CYCLE Next, we investigate how the evolutionary process of metabolic diversification of the primitive biosphere shaped the
planet atmospheric and climatic history. We consider alternate scenarios of biosphere evolutionary complexification (Fig. 4a) consistent with phylogenetic and geological
inferences2,7,9,10,11,27. Our evolutionary sequences culminate with the evolution of anaerobic methanotrophy, which uses the oxidation of CH4 as primary source of energy, based on the
consumption of H2SO4, the main oxidative species of the globally reduced early Earth28. We, therefore, make the plausible assumption that a full methane-cycling biosphere may have evolved
before the advent of photosynthesis. This evolutionary process of diversification may have spanned several hundred million years (from the origin of life 3.9–4.5 Gya, to the origin of
anoxygenic phototrophy, 3.5–3.7 Gya), thus unfolding on a timescale over which the geochemical cycles interacted dynamically and reciprocally with biological activity. In particular, it has
been shown5 that on the timescale of 107 to 108 years, the carbon cycle tends to mitigate temperature variations through a negative feedback on the atmospheric CO2 concentration. We adapted
the model from ref. 5 to add a full, temperature-dependent carbon cycle to our planetary ecosystem model. The abiotic equilibrium _p_CO2 is now determined by the balance between CO2
outgassing and sequestration in the oceanic floor; _p_CH4, by the balance between serpentinization and photodissociation rates; and _p_H2, by the balance between outgassing and photochemical
reactions. Additionally, the H2SO4 oceanic concentration is determined by the balance between rainout and hydrothermal remineralization rates (taken from ref. 29). By drawing values for the
abiotic supplies in CO2, CH4, H2, and H2SO4 from reasonable, log-uniform priors (Table 1), and starting from a lifeless primitive Earth, we evaluate the equilibrium state of the planetary
system after each evolutionary metabolic transition in terms of atmospheric signatures (Fig. 4b–d) and mean surface temperature (Fig. 4e). First, we find that a lifeless Earth is
characterized by a high CO:CH4 atmospheric ratio of 102–104 to 1, which differs markedly from the ratio predicted with a functional biosphere, regardless of its metabolic composition. As the
biosphere complexifies, biological activity increases the atmospheric concentration in CH4, decreases the atmospheric CO, or both, causing the CO:CH4 ratio to fall. By comparing median
values between the abiotic state and the most complex biosphere (MG + AG + AT + MT), the CO:CH4 ratio is predicted to be reduced by a factor of ~5000. The earliest evolutionary events,
whereby the MG, AG, or AG + AT ecosystem emerges, all cause atmospheric shifts that can be distinguished in the _p_CO-_p_H2-_p_CH4 space (Fig. 4b and c). The atmospheric shifts caused by
subsequent evolutionary complexification (leading to MG + AG, MG + AG + AT, or MG + AG + AT + MT ecosystems) are less pronounced and the corresponding atmospheric signatures are less
distinctive among themselves. The evolution of anaerobic methanotrophy (MT) has for instance no effect on the equilibrium atmosphere of the planet because the influx of H2SO4 is sufficient
for methanotrophs to survive and co-occur with methanogens, but not for them to consume a significant portion of the CH4 produced by methanogens. Finally, although methanogenesis can have
major effects on climate on relatively short time scales (Figs. 2 and 3), the carbon cycle buffers these effects at equilibrium on longer time scales: the average temperature difference
between the planet with and without a methanogenic biosphere on its surface is only 4 °C (Fig. 4d). Biological effects on temperature may be further attenuated by the formation of cooling
organic hazes favored by the enhanced sequestration of CO2 in response to methanogenesis25. However, the necessary conditions for organic hazes to form in this case are met in only 0.03% of
the simulations including a methanogenic biosphere. TRANSIENT CLIMATE DESTABILIZATION BY METHANOTROPHY How the planetary atmosphere-climate system responds to metabolic innovation may depend
on the pace of evolution itself. The evolution of methanotrophy is a case in point. Slow evolution may delay methanotrophy after the response of the carbon cycle to methanogenesis. In this
case, the oceanic stocks of H2SO4 are sufficient for methanotrophs to rapidly consume almost all of the atmospheric CH4 (Supplementary Fig. 6) and the planet is temporarily characterized by
an atmospheric deficit in both CO2 and CH4. As a result, temperature plummets below the initial abiotic temperature. In contrast, if evolution is fast enough and methanotrophs evolve before
equilibration of the methanogenic biosphere with the planetary atmosphere and climate, atmospheric CH4 may be consumed during or after the warming period (Fig. 3) but before the deficit in
_p_CO2 builds up; temperature then returns close to its initial value (Supplementary Fig. 6). Figure 5a illustrates the atmospheric and climatic consequences of slow evolution, when the wait
time for methanotrophy is of the order of the carbon cycle timescale. In the two examples shown, the evolution of methanotrophs causes the sharp climatic response described above, and
temperature falls respectively by 8 and 10 °C below _T_Geo = 2 °C, driving the planet into a global glaciation. Among 2000 simulated planetary conditions characterized by randomly drawn
abiotic characteristics (i.e., the abiotic influxes of CO2, H2, CH4 and H2SO4, thereby setting the abiotic _p_CO2, _p_H2, _p_CH4, initial oceanic concentration of H2SO4 and surface
temperature _T_Geo), and evolution time of methanotrophy (from 0 to 125 million years after methanogenesis), 50% experience a temperature drop larger than 8.3 °C below _T_Geo (Fig. 5b), and
40% actually end up in glaciation (Fig. 5b and Supplementary Fig. 7). As expected, delayed evolution of methanotrophy makes extreme cooling more likely (Fig. 5b). Because they lead to an
enhanced abiotic response of the carbon cycle, conditions for which methanogenic ecosystems have the greatest warming effect (low abiotic _T_Geo, high H2 outgassing rate) are also the
conditions under which the evolution of methanotrophy has the most dramatic effect on climate and habitability (Fig. 5a and c, Supplementary Fig. 8). Noticingly, this transient occurs
irrespective of the metabolic composition of the methanogenic biosphere prior to the evolution of methanotrophy, as illustrated in Supplementary Fig. 7. For a methane-cycling ecosystem that
triggers and eventually survives global glaciation, we expect the resulting equilibrium (co-occuring methanogens and methanotrophs with low _p_CH4) to be maintained only transitorily. We
assume that during the early Archean, prior to the evolution of methanotrophy, the oceanic stock of H2SO4 builds up as volcanoes emit SO2 photochemically transformed into H2SO4, which then
deposits on the ocean surface. With a deposition rate corresponding to our lowest value of 107 molecules cm−2 s−1 (ref. 30), and a hydrothermal removal rate of [H2SO4]oc. × 7.2 × 1012 L y−1
(ref. 29), we obtain an abiotic oceanic concentration of 0.4 mM. Such a stock is sufficient for methanotrophs to consume most of the atmospheric CH4, leading to the global cooling described
above. Since the rate of CH4 reduction by methanotrophs is faster than the deposition rate, the H2SO4 stock will ultimately be depleted by methanotrophs, thereby driving a new abrupt
environmental shift towards the equilibrium described in Fig. 4. At this new, stable equilibrium of coexisting methanogens and methanotrophs, the atmospheric _p_CH4 is high and the H2SO4
oceanic concentration is very low, below the micromolar, which aligns with the sulfur isotopic fractionation records30,31,32. This result suggests that anaerobic methanotrophy may have been
the main sink of H2SO4 prior to the Neoarchean (2.5–2.7 Gya). DISCUSSION Our initial questions were: How constrained was the habitability of late Hadean/early Archean Earth to
methane-cycling ecosystems? How productive were these ecosystems and did they have a significant impact on the atmospheric composition and climate? How did their impact change as different
metabolisms evolved? To address these questions, we build on the previous theory8,12,13 by setting up a probabilistic modeling framework in which the evolution of a microbial biosphere is
coupled to the dynamics of early Earth surface geochemistry and climate. We focus on four simple microbial metabolisms involved in methane cycling and likely to be some of the earliest
players in Earth’s ecology: hydrogenotrophic methanogenesis (MG), acetogenesis (AG), methanogenic acetotrophy (AT), and anaerobic oxidation of methane (MT). The biosphere evolves when a
metabolism that was not present appears and adapts. By closing the global feedback loop between biological and planetary surface processes, the model predicts both ecosystem and
atmosphere-climate states under the assumption that they reciprocally influence one another. Our results confirm the contention that the late Hadean/early Archean planet was most likely
habitable to methane-cycling chemolithotrophic biospheres and that under the assumption of high enough H2 supply, these biospheres were key factors of the climatic and atmospheric evolution
of the planet8,12,13,33,34. On short time scales (105–106 years) the evolution of methanogenic biospheres may have considerably warmed the climate and influenced its resilience, in spite of
a very low ecosystem productivity. On longer timescales, commensurate with the abiotic response of the carbon cycle to temperature variation, all ecosystems converge to new stable
equilibria. Under these long-term equilibrium conditions, the mean surface temperature does not differ much from the lifeless state, and the carbon cycle is the predominant mechanism of
climate regulation. However, all ecosystem equilibria share a robust atmospheric signature (low CO:CH4 ratio), distinctive from the lifeless state. In addition to influencing planetary
characteristics at equilibrium, metabolic evolution generates atmospheric and climatic transients. The pace of evolution thus has a strong influence on the atmosphere and climate history. In
particular, fast evolution of methanotrophs has limited or no effect on climate, whereas their delayed evolution may cause strong transients leading to global glaciation. Although the
influence of chemotrophic methane-cycling ecosystems on climate has been discussed in the previous work8,12,13,33,34, our model is the first to couple models of the Archean atmosphere,
climate, and carbon cycle to an explicit eco-evolutionary model of cell population dynamics in order to quantify this effect. Our model differs from previous work primarily by addressing how
climate change driven by ecological function feeds back to the biological activity of microbial populations, through the thermal dependence of the thermodynamics and kinetics of cell
metabolism, and triggers an abiotic response of the carbon cycle. Closing the global feedback loop between ecological and planetary processes allows us to predict the ecosystem and climate
states under the assumption that they reciprocally influence one another on multiple timescales. A general result is that MG and AG + AT ecosystems are characterized by extremely low biomass
production relative to their planetary impact on the atmosphere and climate. We predict biomass production to be 1–4 orders of magnitude smaller than previous estimates13 and 3–7 orders of
magnitude below modern values26. Maximum global biomass production (reached for a very specific combination of biotic and abiotic conditions) is 1010 molecules C cm−2 s−1, or 3 × 107 ton C
year−1. This is ~1000 times less than estimates of modern primary production. Both in terms of stock and fluxes, biomass has therefore very little effect on the biogeological coupling, which
is a major difference with modern ecosystems. A key assumption of our model is that nutrients N and P are not limiting; this assumption is backed up by the prediction of very low biomass
production. As previously argued13,14, primitive ecosystems with such low productivity should have been limited by the availability in electron donors rather than by N and P nutrients. More
specifically, ref. 14 evaluated the most likely limitation to biomass production prior to the evolution of oxygenic photosynthesis. By assuming a fixed yield for biomass production per
molecules of electron donor consumed (as in ref. 13), they found that the N and P supplies were most likely sufficient during the Archean for electron donors to be limiting. This is even
more likely given our finding of a biomass productivity far lower than the previous estimates13,14. Our prediction of very low biomass productivity may help better constrain the timeline of
the evolution of metabolic innovation on Earth. Even in our most productive scenario and under the (extreme) assumption that 100% of the dead organic matter is buried before
remineralization, the estimated biomass production is still 4–5 times lower than the level consistent with the C isotopic fractionation estimated from rocks as old as 3.5 Gy1,35. This
suggests that more productive, likely photosynthetic life forms must have evolved more than 3.5 Gya ago. By performing equilibrium analyses of the planetary system on longer time-scales
(107–108 years), on which the carbon cycle responds to ecosystem function and sequential metabolic diversification of the biosphere, we find that the climate regulation of the planet by the
abiotic carbon cycle largely buffers the influence of early methanogenic activity on climate. However, depending on the pace of evolution, metabolic transitions such as the evolution of
methanotrophy can interact with the abiotic carbon cycle and trigger strong transitory climatic events such as global glaciation over 107–108 years. Even though the timing of the origin of
anaerobic oxidation of methane (AOM), by reduction of nitrate, nitrite, or sulfate, is not well resolved, the fact that AOM proceeds enzymatically as a reversal of methanogenesis has been
used to suggest that AOM may have evolved relatively soon after methanogens, i.e. before phototrophy36,37. Additionally, we find that the evolution of AOM prior to photosynthesis appears to
be compatible with the sulfur isotopic fractionation record30,31,32. Our results thus highlight biological activity and the evolution of pre-photosynthetic methane-cycling ecosystems as a
potential destabilizing factor of the early Earth climate system, alternatively or additionally to abiotic causes such as large impactors3,4,38. The mechanism by which the evolution of early
methane-cycling ecosystems may have exposed the Earth to high risks of global glaciation—fast removal of atmospheric methane, slow response of the carbon cycle—is general. It is, therefore,
tantalizing to speculate about its potential involvement in the major climatic events that paralleled the evolution of oxygenic photosynthesis. By poisoning methanogens, removing the
resource limitation (oxidizing species: O2, H2SO4, NO3, NO2) of methanotrophs, or by simply driving the abiotic oxidation of atmospheric CH4 by outgassed O2, oxygenic photosynthesis may have
driven a dramatic decline in atmospheric CH4 which, coupled to a delayed response of the carbon cycle, could have caused global cooling, triggering the Proterozoic glaciation ~2.3
Gya39,40,41. The substantial shifts in atmospheric composition driven by evolutionary metabolic innovation (Fig. 4) highlight the fact that simple methane-cycling ecosystems can have
radically different atmospheric signatures. Thus, the atmosphere composition is shaped very early on by the course of biological evolution. We observe that the atmospheric signature of a
specific metabolism is highly dependent on the ecological context set by the whole metabolic composition of the biosphere. As the biosphere evolves and complexifies, the atmospheric
signature of each metabolic type becomes less identifiable, yet the state of the planet remains distinctive from the lifeless scenario. This backs up and quantifies the hypothesis that on a
globally reduced planet such as the early Archean Earth, a low CO:CH4 atmospheric ratio is a highly discriminant indicator of inhabitation by a primitive CH4-based biosphere42. In
conclusion, our results suggest that life has had a dramatic impact on the anoxic Earth’s surface environment, long before phototrophy evolved. They highlight the importance of the
evolutionary process and its timeline in shaping up the planet’s atmosphere and climate. In spite of extremely low productivity, metabolic evolutionary innovation in primitive methane-based
biospheres is predicted to cause distinctive shifts in atmospheric composition, such as a decreasing CO:CH4 ratio as greater metabolic complexity evolves. The warming effect of methanogens
and cooling effect of methanotrophs can be strong but they are only transient, on timescales that depend on the pace of evolution. We anticipate that the continued development of models that
couple planetary processes with ecological and evolutionary dynamics of microbial biospheres will further advance our understanding of major events in the co-evolutionary history of life
and Earth, and help identify detectable biosignatures for the search of life on Earth-like exoplanets. METHODS BIOLOGICAL MODEL In the following section, we present the biological model.
Parameters’ definition, unit and default value are given in Supplementary Tables 1, 2, and 3. The biological model describes the dynamics of one or several biological populations of
chemotrophic organisms, driven by the growth, birth (division) and death of individual cells. The individual cell life cycle is controlled by catabolic and anabolic reactions occurring
within cells17,18,43. Catabolism produces energy used by anabolism for biomass production, which determines cell growth and division; a fraction of energy produced by catabolism is used for
cell maintenance. Energy thus flows from catabolism to maintenance and anabolism, and these processes can be described as follows: $$\begin{array}{*{20}{l}} {{\mathrm{Catabolism}}} \hfill
& : \hfill & {\mathop {\sum }\limits_{i = 1}^n \gamma _{S_i}^{cat}S_i \to \mathop {\sum }\limits_{i = 1}^m \gamma _{P_i}^{cat}P_i + {\mathrm{\Delta }}G_{cat}} \hfill \\
{{\mathrm{Maintenance}}} \hfill & : \hfill & {E_m} \hfill \\ {{\mathrm{Anabolism}}} \hfill & : \hfill & {\mathop {\sum }\limits_{i = 1}^{\dot n} \gamma _{S_i}^{cat}S_i +
{\mathrm{\Delta }}G_{ana} \to \mathop {\sum }\limits_{i = 1}^{\dot m} \gamma _{P_i}^{cat}P_i + B} \hfill \end{array}$$ (E1) where _S__i_ and _P__i_ are the substrates and products of the
metabolic reactions, and the \(\gamma\)’s are the corresponding stoichiometric coefficients. We follow ref. 17 and assume that the average organic compound
\({\mathrm{C}}{\mathrm{H}}_{1.8}{\mathrm{O}}_{0.5}{\mathrm{N}}_{0.2}\) is a plausible approximation for the living biomass _B_. The energy _E__m_ (in kJ d−1) measures the cost of maintenance
for a single cell per unit of time. The ∆_G_ terms measure the oxidative power released or consumed by the catabolic and anabolic reactions; they are given by the Nernst relationship:
$${\mathrm{\Delta }}G(T) = {\mathrm{\Delta }}G_0(T) + {RTlog} \left(\mathop {\prod }\limits_{i = 1}^n S_i^{ \gamma _{S_i}}\mathop {\prod }\limits_{i = 1}^m P_i^{\gamma _{P_i}}\right)$$ (E2)
where _R_ is the ideal gas constant, _T_ is the temperature (in K) and ∆_G_0(_T_) (in kJ) is the Gibbs energy of the reaction. ∆_G_0(_T_) is obtained from the Gibbs–Helmholtz relationship:
$${\mathrm{\Delta }}G(T) = {\mathrm{\Delta }}G_0(T_S)\frac{T}{{T_S}} + {\mathrm{\Delta }}H_0(T_S)\frac{{T_S - T}}{{T_S}}$$ (E3) where _T__S_ is the standard temperature of 298.15 K. Note
that equations (E2) and (E3) describes how the thermodynamics of any given metabolism (i.e., the combination of catabolism and anabolism) vary with temperature. Michaelis–Menten kinetics
apply to catabolism: $$q_{cat} = q_{max}\frac{{min(S_i^{cat}/\gamma _{S_i}^{cat})}}{{min(S_i^{cat}/\gamma _{S_i}^{cat}) + K_S}}$$ (E4) where _K__S_ is the half-saturation constant, _q__max_
the maximum metabolic rate of the cell (_d_−1), and \(min(S_i^{cat}/\gamma _{S_i}^{cat})\) measures the concentration of the limiting substrate (taking stoichiometry into account). The
energy produced is first directed toward maintenance. The cell energetic requirement for maintenance is: $$q_m = \frac{{ - E_m}}{{{\mathrm{\Delta }}G_{cat}}}$$ (E5) The cell therefore meets
its energy requirement only if _q__cat_ > _q__m_. If the cell does not meet this requirement, the basal mortality rate of the cell, _m_, (in _d_−1), is augmented by a decay-related
mortality term equal to \(k_d(q_m - q_{cat})\). Hence the actual mortality rate: $$\begin{array}{*{20}{l}} {If\,q_{cat} \,<\, q_m} \hfill & : \hfill & {d = m + k_d(q_m - q_{cat})}
\hfill \\ {If\,q_{cat} \,> \, q_m} \hfill & : \hfill & {d = m} \hfill \end{array}$$ (E6) When the energy requirements for maintenance are not met (_q__cat_ < _q__m_) no energy
is allocated to biomass production. Conversely, when those requirements are met (_q__cat_ < _q__m_), then the energy remaining after allocation to cell maintenance can be directed to
anabolism. A constant quantity of energy\({\mathrm{\Delta }}G_{diss}\) (in kJ) is then lost through dissipation. Following ref. 17 we consider the following empirical relationship:
$${\mathrm{\Delta }}G_{diss} = 200 + 18(6 - NoC)^{1.8} + e^{( - 0.2 - \alpha )^{2.16}(3.6 + 0.4NoC)}$$ (E7) where _NoC_ is the number of carbons in the carbon source used by the anabolic
reaction, and \(\alpha\) is its degree of oxidation. The efficiency of metabolic coupling (i.e., the number of occurrences of the catabolic reaction to fuel one occurrence of the anabolic
reaction once the maintenance cost has been met) is then measured by $$\lambda = \frac{{ - {\mathrm{\Delta }}G_{cat}}}{{{\mathrm{\Delta }}G_{ana} + {\mathrm{\Delta }}G_{diss}}}$$ (E) The
Michaelis–Menten kinetics of anabolism is given by $$\begin{array}{*{20}{l}} {If\,q_{cat} \,> \, q_m} \hfill & : \hfill & {q_{ana} = \lambda (q_{cat} -
q_{m})\frac{{min(S_i^{ana}/\gamma _{S_i}^{ana})}}{{min(S_i^{ana}/\gamma _{S_i}^{ana}) + K_S}}} \hfill \\ {If\,q_{cat} \,<\, q_m} \hfill & : \hfill & {q_{ana} = 0} \hfill
\end{array}$$ (E9) where the half-saturation constant _K__S_ independent of the substrate, as was assumed in ref. 43. As biomass accumulates in the cell at rate _q__ana_, cell growth may
lead to cell division. The cell division rate, _r_, is given by: $$\begin{array}{*{20}{l}} {If\,B \,<\, 2B_{Struct}} \hfill & : \hfill & {r = 0} \hfill \\ {If\,B \,> \,
2B_{Struct}} \hfill & : \hfill & {r = r_{max}\frac{1}{{1 + e^{ - \theta log10((B - 2B_{Struct})/B_{Struct})}}}} \hfill \end{array}$$ (E10) This means that a cell cannot divide if its
internal biomass is not at least twice its cell structural biomass so that the two daughter cells meet this structural requirement. Above this threshold value of 2_B__Struct_, _r_ is of
sigmoidal form so that the division rate first increases exponentially with the intracellular biomass content then saturates to _r__max_ when biomass is largely available. Using the rates of
catabolic and anabolic cell activity and resulting cell division and mortality rates, we derive the following system of ordinary differential equations driving the cell population dynamics
(dynamics of the number of cells and average cell biomass) and the feedback of the population on its environment (chemical composition of the ocean surface): $$\begin{array}{*{20}{l}}
{\frac{{dN_i}}{{dt}}} \hfill & = \hfill & {(r_i - d_i)N_i} \hfill \\ {\frac{{dB_i}}{{dt}}} \hfill & = \hfill & {q_{ana_i} - r_iB} \hfill \\ {\frac{{dX_j}}{{dt}}} \hfill &
= \hfill & {F(X_j) + \mathop {\sum }\limits_{i = 1}^{MT} (q_{cat_i}\gamma _{i,X_j}^{cat} + q_{ana_i}\gamma _{i,X_j}^{ana})N_i} \hfill \end{array}$$ (E11) where _MT_ denotes the ensemble
of metabolic types considered, _N__i_ is the number of individual cells in a population of a given metabolic type, _B__i_ is the average cellular biomass of that type, and \(X_1,...,X_S\)
are the concentrations of all relevant chemical species in the environment. The term \({\sum }_{i = 1}^{MT} (q_{cat_i}\,\gamma _{i,X_j}^{cat} + q_{ana_i}\gamma _{i,X_j}^{ana})N_i\) describes
how concentrations vary according to the biological activity of each biological population _i_ in the microbial community. The _F_(_X__j_) terms describe the environmental forcing resulting
from ocean circulation and atmosphere-ocean exchanges as simulated by a stagnant boundary layer model as in ref. 13. The system of equations (E11) is solved numerically using a forward
Euler method. The flow of energy through the cell is driven by the maximum metabolic rate, _q__max_ (in _d_−1), and the rate of energy consumption for maintenance, _E__m_ (in kJ d−1). They
are both expressed as functions of temperature and cell size of the form _e__a_+_b__T_Vc. Default values for parameters _a_, _b_ and _c_ entering _q__max_ and _E__m_ are given in
Supplementary Table 2. The structural cell biomass _B__Struct_ increases with cell size according to _B__Struct_ = _aV__b_. Both metabolic rate and maintenance cost increase with cell size,
but not as fast as structural biomass. Consequently, the biomass specific rates of metabolism and energy consumption for maintenance decrease with cell size. Thus, small organisms are
better at acquiring energy, but large organisms are more cost efficient due to lower maintenance requirements. A trade-off mediated by cell size thus exists between metabolic and maintenance
rates, hence an optimal (intermediate) cell size, which is consistent with previous work conducted on unicellular marine organisms22. Both metabolic and maintenance rates increase with
temperature, but the metabolic rate increases slightly faster19,20, shifting the trade-off toward larger sizes. As a consequence, the optimal cell size increases with temperature. To compute
the evolutionarily optimal cell size for a given metabolic type at a given temperature, we run simulations across a range of cell size and measure the level of resource use at equilibrium
given by \(Q^ \ast = \mathop{\prod}\limits_{i = 1}^n {S_i^{ \gamma _{S_i}^{cat}}} \mathop{\prod}\limits_{i = 1}^m {P_i^{\gamma _{P_i}^{cat}}}\), the product of the metabolic substrates and
wastes concentrations at equilibrium weighted by their stoichiometric coefficient. According to the classical principle of “pessimization” (maximization of resource use)44,45,46, populations
that are better at exploiting their environment have lower _Q_* and evolution by natural selection will favor the cell size that minimizes _Q_*, thus leading to the evolutionarily optimal
cell size. Supplementary Fig. 9 shows _Q_* as a function of cell size and temperature for the H2-based methanogenesis. The optimal cell size, _S__C_*, follows a positive relationship with
temperature given by _S__C_* = 10_a_+_bT_. We consider that over geological time scales, adaptive evolution acting on genetic variation among cells is fast enough so that _S__C_ is equal to
_S__C_*. As temperature changes, evolutionary adaptation by natural selection tracks the temperature-dependent optimal cell size. In contemporary Earth oceans, most of the dead biomass is
recycled by fermentors (>99%), and the rest is buried into the ocean floor. Although the most general version of our model includes populations of acetogenic biomass fermentors and
acetotrophs, their inclusion considerably increases simulation time (results not shown). All the results presented in the main text have been obtained without fermentors, assuming that the
dead biomass accumulates in the ocean’s interior. It appears that biomass productivity is so low that the effect of biomass recycling (or lack thereof) on the atmospheric composition is
negligible. We verified this by testing the effect of a recycling pathway in each ecosystem (MG, AG + AT, MG + AG + AT), for an intermediate value of H2 volcanic outgassing. Although biomass
production is sufficient to sustain a population of fermentors, we found no significant effect of their biological activity on the atmospheric composition. Additionally, when we can compare
the global atmospheric redox budget of the planet with and without biomass production (see Supplementary Results and Discussion, Supplementary Fig. 12), we find no significant differences.
This further demonstrates that biomass production and the accumulation of dead biomass in the absence of remineralization represents a sink of C and H that is marginal compared to the other
fluxes in the model. Note that the situation would be very different after the evolution of more productive, photosynthetic primary producers. Because we do not model the fate of dead
biomass explicitly, we evaluate the consistency of our predictions of biomass production with the geological record (carbon isotopic fractionation data) by taking biomass production as the
upper bound for burial (in which case 100% of the produced biomass is ultimately buried) and using the modern value of burial (taken to be 0.2% as in e.g., ref. 13) as lower bound. PLANETARY
MODEL Our computation of climate state and mean surface temperature is based on 3D simulations with the Generic LMD GCM4,47,48. The model includes a 2-layer dynamic ocean computing heat
transport and sea ice formation49. The radiative transfer is based on the correlated-_k_ methods with _k_-coefficients calculated using the HITRAN 2008 molecular database. We used the
simulations described in ref. 4 for the early Earth at 3.8 Ga, assuming no land, 1 bar of N2 and a 14 h rotation period. The simulation grid covers a range of _p_CO2 from 0.01 to 1 bar and
pCH4 from 0 to 10 mbar. From this grid, we derived the following simple parametrization of the mean surface temperature as a function of _p_CO2 and _p_CH4 (expressed in bar): $$T\left( {\!
\deg {\mathrm{C}}} \right) = - 19.26 + 77.67 {\sqrt {p{\mathrm{C}}{{\mathrm{O}}_2}}} \,\,+\, 5{\mathrm{log}}10\left( {\frac{{1 + p{\mathrm{C}}{{\mathrm{H}}_4}{{10}^6}}}{3}} \right)$$ (E12)
In the coupled biological-planetary model, we assume that the climate is always at equilibrium, meaning that the timescale of climate convergence is shorter than biological and geochemical
timescales. Photochemistry is computed with the 1D version of the Generic LMD GCM, which now includes a photochemistry core from ref. 50. The chemical network includes 30 species (mostly
hydrocarbons) and 114 reactions. We use the reactions from ref. 50 for H2-H2O-CO2 and from ref. 51 for hydrocarbons. The photochemistry model also includes a pathway for the formation of
hydrocarbon aerosol (C2H + C2H2 → HCAER + H)51,52. We use the eddy diffusion vertical profile from ref. 53 and the solar UV spectrum at 3.8 Ga from ref. 54. Boundary conditions are set by
the mixing ratios of CO2, CH4, and H2O at the first atmospheric layer. The atmospheric CO is either consumed biotically by acetogens when they are present in the biosphere, or abiotically by
the formation of formate in the ocean (resulting in a deposition velocity of ~108 cm s−1) as in ref. 13,55. This eliminates the possibility of CO-runaway, which otherwise occurs in our
simulations at high level of H2 or CO2. In addition, we assume that H2 undergoes diffusion-limited atmospheric escape (see for instance ref. 13). We performed simulations across a range of
_p_CO2 (104–105 ppm), _p_CH4 (1–104 ppm), H2 (100–104 ppm). For our range of atmospheric composition, photochemistry is dominated by the photolysis of CO2 and CH4, whose net reactions are:
$${\mathrm{CO}}_2 + {\mathrm{H}}_2 = {\mathrm{CO}} + {\mathrm{H}}_2{\mathrm{O}}$$ (R1) $${\mathrm{CH}}_4 + {\mathrm{CO}}_2 = 2{\mathrm{CO}} + 2{\mathrm{H}}_2$$ (R2) From the simulation
outputs we derived simple parametrizations for the production rates and loss rates of CH4, CO, H2 (see Supplementary Fig. 10). The reaction rates of (R1) and (R2) can be parameterized
respectively as (in molecules cm−2 s−1): $$F_1 = 1.8\,10^{10} \cdot \left( {\frac{{p{\mathrm{C}}{\mathrm{O}}_2}}{{10^{ - 1}}}} \right) \cdot \left( {\frac{{p{\mathrm{H}}_2}}{{10^{ - 4}}}}
\right)^{0.4}$$ (E13) $$F_2 = 2\,10^{10}\left( {\frac{{p{\mathrm{C}}{\mathrm{O}}_2}}{{10^{ - 1}}}} \right)^{ - 0.2} \,\cdot\, \left( {\frac{{p{\mathrm{H}}_2}}{{10^{ - 4}}}} \right)^{ - 0.2}
\,\cdot\, \left( {\frac{{p{\mathrm{C}}{\mathrm{H}}_4}}{{10^{ - 4}}}} \right)^u$$ (E14) where _u_ (between 0.5 and 1) is given by $$u = 0.6 + 0.1{\it{{\mathrm{log}}}}\left(
{\frac{{p{\mathrm{H}}_2}}{{10^{ - 4}}}} \right) - \frac{{{\it{{\mathrm{log}}}}({\it{p{\mathrm{C}}{\mathrm{H}}}}_4) + 2}}{{15}} + 0.08\log \left(\frac{{p{\mathrm{C}}{\mathrm{O}}_2}}{{10^{ -
1}}}\right)$$ (E15) To simulate the evolution of _p_CO2 with time and feedbacks between the microbial community, climate, and the carbon cycle, we use a simple carbon cycle model based on
ref. 5. The model computes the evolution of atmospheric CO2, dissolved inorganic carbon (CO2, HCO3− and CO32−) and pH in the ocean and in seafloor pores. The model takes into account
outgassing (from volcanoes and mid-oceanic ridges), continental silicate weathering, dissolution of basalts in the seafloor, and oceanic chemistry, as sources and sinks of CO2. For all
parameters in the carbon cycle model, we use the mean values given in ref. 5. For the temperature dependences of silicate weathering and oceanic chemistry, we use the mean surface
temperature from our climate model. Even though this model only computes global mean quantities and does not take into account the organic matter in carbon sources and sinks of carbon, it is
computationally very fast and well suited for studying the major feedbacks between biological populations and the atmosphere and climate. COUPLED BIOLOGICAL-PLANETARY MODEL The model
resolves dynamics that take place on extremely different timescales—geological processes are extremely slow (103–106 years) compared to biological dynamics (days to years). We use a
timescale separation approach and assume that the planetary environment is fixed on the biological time scale, and the biological system (population and local environment) is at ecological
and evolutionary equilibrium on the geological time scale. This allows us to resolve the biological and geological dynamics separately and couple them at discrete points in time. The
biological dynamics are first resolved by the biological model for a fixed environmental forcing (equations (E11)), with microbial cells at their evolutionarily optimal size. The biological
model is integrated over a sufficiently long time so that the local ecosystem reaches its equilibrium. Then we compute the biogenic fluxes between the surface ocean and the atmosphere, and
between the surface ocean and the deep ocean, and feed them into the planetary model. The planetary model is then used to resolve the system’s geochemical and climate dynamics, but only for
a sufficiently small change in the planetary state (1% to 1‰ variation of the state variable that changes the most) so that this change would alter the biological equilibrium only
marginally. The new biological equilibrium is then computed, and the process is re-iterated. This iterative process approximates a continuous coupling between the microbial community and its
planetary environment. MONTE-CARLO SIMULATIONS We explore the parameter range by stochastically varying the parameters that determine the maximum metabolic rate, the rate of energy
consumption for maintenance, the metabolic half-saturation constant, the decay and mortality rates, and the maximum division rate. We also vary the values of the parameters that shape the
dependencies on size, temperature, and cellular biomass (see Supplementary Table 4). Parameter values are picked uniformly or log-uniformly (to avoid making prior assumptions on the
likelihood of specific regions of the parameter space) within ranges constrained by empirical data from the literature, or large enough to cover plausible empirical variation. Three thousand
simulations were run for the MG ecosystem, one thousand for the AG + AT ecosystem, and one thousand for the MG + AG + AT ecosystem, under four different values of volcanic outgassing:
\(\phi _{{\mathrm{Volc}}}({\mathrm{H}}_2)\) = 2 × 109.5, 2 × 1010, 2 × 1010.5, and 2 × 1011 molecules cm−2 s−1, hence a total of 20,000 simulations. Each simulation was run to equilibrium.
The results can be divided into two subsets of roughly equivalent size, regardless of the ecosystems and volcanic activity considered: those with biological activity and those without
(Supplementary Fig. 1A, B). We then verified that the number of simulations run for each of the considered scenarios was sufficient for the resulting distribution of equilibrium states to
have converged. We evaluated convergence by subsampling, for each scenario, the subset of ‘biologically viable’ simulations with increasing sample size, and computing the average equilibrium
biomass and CH4 biogenic emission of the subsamples. The results are shown in Supplementary Fig. 2 for the three scenarios and for \(\phi _{{\mathrm{Volc}}}({\mathrm{H}}_2)\) = 2 × 1010.5
molecules cm−2 s−1. DATA AVAILABILITY The data used to produce all the results presented in this study are available upon reasonable request to B.S. CODE AVAILABILITY The codes used in this
study to produce the data analyzed are available on a git repository upon reasonable request to B.S. REFERENCES * Nisbet, E. G. & Sleep., N. H. The habitat and nature of early life.
_Nature_ 409, 1083 (2001). Article ADS CAS PubMed Google Scholar * Martin, W. F. & Sousa, F. L. Early microbial evolution: the age of anaerobes._Cold Spring Harb. Perspect. Biol._
8, a018127 (2016). Article PubMed Central CAS Google Scholar * Pearce, B. K. D. et al. Constraining the time interval for the origin of life on Earth. _Astrobiology_ 18, 343–364 (2018).
Article ADS CAS PubMed Google Scholar * Charnay, B. et al. A warm or a cold early Earth? New insights from a 3-D climate-carbon model. _Earth Planet. Sci. Lett._ 474, 97–109 (2017).
Article ADS CAS Google Scholar * Krissansen-Totton, J. et al. Constraining the climate and ocean pH of the early Earth with a geological carbon cycle model. _Proc. Natl Acad. Sci. USA_
115, 4105–4110 (2018). Article ADS CAS Google Scholar * Catling, D. C. & Kasting, J. F. _Atmospheric Evolution on Inhabited and Lifeless Worlds_ (Cambridge University Press, 2017). *
Battistuzzi, F. U. et al. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land._BMC Evol. Biol._ 4, 44 (2004).
Article PubMed PubMed Central CAS Google Scholar * Ozaki, K. et al. Effects of primitive photosynthesis on Earth’s early climate system. _Nat. Geosci._ 11, 55 (2018). Article ADS CAS
Google Scholar * Weiss, M. C. et al. The physiology and habitat of the last universal common ancestor. _Nat. Microbiol._ 1, 16116 (2016). Article CAS PubMed Google Scholar * Marin, J.
et al. The timetree of prokaryotes: new insights into their evolution and speciation. _Mol Biol Evol._ 34, 437–446 (2016). Google Scholar * Havig, J. R. et al. Sulfur and carbon isotopic
evidence for metabolic pathway evolution and a four-stepped Earth system progression across the Archean and Paleoproterozoic. _Earth-Sci. Rev._ 174, 1–21 (2017). Article ADS CAS Google
Scholar * Kasting, J. F. et al. A coupled ecosystem-climate model for predicting the methane concentration in the Archean atmosphere. _Orig. Life Evol. Biosph._ 31, 271–285 (2001). Article
ADS CAS PubMed Google Scholar * Kharecha, P., Kasting, J. & Siefert, J. A coupled atmosphere–ecosystem model of the early Archean Earth. _Geobiology_ 3, 53–76 (2005). Article CAS
Google Scholar * Ward, L. M., Rasmussen, B. & Fischer, W. W. Primary productivity was limited by electron donors prior to the advent of oxygenic photosynthesis. _J. GeoPhys. Res.
BioGeo._ 124, 211–226 (2019). * Charnay, B. et al. Exploring the faint young Sun problem and the possible climates of the Archean Earth with a 3‐D GCM. _J. Geophys. Res. Atmos._ 118, 10–414
(2013). Article CAS Google Scholar * Lefèvre, F. et al. Three‐dimensional modeling of ozone on Mars. _J. Geophys. Res. Planets_ 109, E7 (2004). Article CAS Google Scholar *
Kleerebezem, R. & Van Loosdrecht., M. C. M. A generalized method for thermodynamic state analysis of environmental systems. _Crit. Rev. Environ. Sci. Tec._ 40, 1–54 (2010). Article
Google Scholar * González-Cabaleiro, R., Lema, J. M. & Rodríguez, J. Metabolic energy-based modelling explains product yielding in anaerobic mixed culture fermentations._PLoS ONE_ 10,
e0126739 (2015). Article PubMed PubMed Central CAS Google Scholar * Tijhuis, L., Van Loosdrecht, M. C. M. & Heijnen, J. J. A thermodynamically based correlation for maintenance
Gibbs energy requirements in aerobic and anaerobic chemotrophic growth. _Biotechnol. Bioeng._ 42, 509–519 (1993). Article CAS PubMed Google Scholar * Gillooly, J. F. et al. Effects of
size and temperature on metabolic rate. _Science_ 293, 2248–2251 (2001). Article ADS CAS PubMed Google Scholar * Litchman, E. et al. The role of functional traits and trade‐offs in
structuring phytoplankton communities: scaling from cellular to ecosystem level. _Ecol. Lett._ 10, 1170–1181 (2007). Article PubMed Google Scholar * Ward, B. A. et al. The size dependence
of phytoplankton growth rates: a trade-off between nutrient uptake and metabolism. _Am. Nat._ 189, 170–177 (2017). Article PubMed Google Scholar * Aksnes, D. L. & Egge, J. K. A
theoretical model for nutrient uptake in phytoplankton. _Marine Ecol. Prog. Ser. Oldendorf_ 70, 65–72 (1991). Article ADS Google Scholar * Arney, G. et al. The pale orange dot: the
spectrum and habitability of hazy Archean Earth. _Astrobiology_ 16, 873–899 (2016). Article ADS CAS PubMed PubMed Central Google Scholar * Domagal-Goldman, S. D. et al. Organic haze,
glaciations and multiple sulfur isotopes in the Mid-Archean Era. _Earth Planet. Sci. Lett._ 269, 29–40 (2008). Article ADS CAS Google Scholar * Prentice, I. C. et al. _The Carbon Cycle
and Atmospheric Carbon Dioxide_, 185–237 (Cambridge University Press, 2001). * Moore, E. K. et al. Metal availability and the expanding network of microbial metabolisms in the Archaean eon.
_Nat. Geosci._ 10, 629–636 (2017). Article ADS CAS Google Scholar * Catling, D. C., Claire, M. W. & Zahnle, K. J. Anaerobic methanotrophy and the rise of atmospheric oxygen. _Philos.
Trans. A Math. Phys. Eng. Sci._ 365, 1867–1888 (2007). Article ADS CAS PubMed Google Scholar * Wong, M. et al. Nitrogen oxides in early Earth’s atmosphere as electron acceptors for
life’s emergence. _Astrobiology_ 17, 975–983 (2017). Article ADS CAS PubMed Google Scholar * Ono, S. et al. New insights into Archean sulfur cycle from mass-independent sulfur isotope
records from the Hamersley Basin, Australia. _Earth Planet. Sci. Lett._ 213, 15–30 (2003). Article ADS CAS Google Scholar * Habicht, K. S. et al. Calibration of sulfate levels in the
Archean ocean. _Science_ 298, 2372–2374 (2002). Article ADS CAS PubMed Google Scholar * Crowe, S. A. et al. Sulfate was a trace constituent of Archean seawater. _Science_ 346, 735–739
(2014). Article ADS CAS PubMed Google Scholar * Kasting, J. F. Methane and climate during the Precambrian era. _Precambrian Res._ 137, 119–129 (2005). Article ADS CAS Google Scholar
* Kasting, J. F. & Ono, S. Palaeoclimates: the first two billion years. _Philos. Trans. R Soc. Lond. B Biol. Sci._ 361, 917–929 (2006). Article CAS PubMed PubMed Central Google
Scholar * Krissansen-Totton, J. et al. A statistical analysis of the carbon isotope record from the Archean to Phanerozoic and implications for the rise of oxygen. _Am. J. Sci._ 315,
275–316 (2015). Article ADS CAS Google Scholar * Knittel, K. & Boetius, A. Anaerobic oxidation of methane: progress with an unknown process._Ann. Revi. Microbiol._ 63, 311–334
(2009). Article CAS Google Scholar * Nitschke, W. & Russell, M. J. Beating the acetyl coenzyme A-pathway to the origin of life. _Philos. Trans. R Soc. Lond. B Biol. Sci._ 368,
20120258 (2013). Article PubMed PubMed Central CAS Google Scholar * Sleep, N. H. & Zahnle, K. Carbon dioxide cycling and implications for climate on ancient Earth. _J. Geophys. Res.
Planets_ 106, 1373–1399 (2001). Article ADS CAS Google Scholar * Laakso, T. A. & Schrag, D. P. Methane in the Precambrian atmosphere. _Earth Planet. Sci. Lett._ 522, 48–54 (2019).
Article ADS CAS Google Scholar * Lyons, T. W., Reinhard, C. T. & Planavsky, N. J. The rise of oxygen in Earth’s early ocean and atmosphere. _Nature_ 506, 307 (2014). Article ADS
CAS PubMed Google Scholar * Kopp, R. E. et al. The Paleoproterozoic snowball Earth: a climate disaster triggered by the evolution of oxygenic photosynthesis. _Proc. Natl Acad. Sci. USA_
102, 11131–11136 (2005). Article ADS CAS PubMed Google Scholar * Krissansen-Totton, J., Olson, S. & Catling, D. C. Disequilibrium biosignatures over Earth history and implications
for detecting exoplanet life. _Sci. Adv._ 4, eaao5747 (2018). Article ADS PubMed PubMed Central Google Scholar * González-Cabaleiro, R. et al. Microbial catabolic activities are
naturally selected by metabolic energy harvest rate. _ISME J._ 9, 2630 (2015). Article PubMed PubMed Central CAS Google Scholar * Tilman, D. _Resource Competition and Community
Structure_ (Princeton University Press, 1982). * Mylius, S. D. & Diekmann, O. On evolutionarily stable life histories, optimization and the need to be specific about density dependence.
_Oikos_ 74, 218–224 (1995). * Metz, J. A. J., Mylius, S. D. & Diekmann, O. When does evolution optimise? _Evol. Ecol. Res._ 10, 629–654 (2008). * Wordsworth, R. et al. Modelling past
Mars Climates and water cycle with a thicker CO2 atmosphere. _Mars Atmosphere: Modelling and Observation_ 447–448 (2011). * Leconte, J. et al. Increased insolation threshold for runaway
greenhouse processes on Earth-like planets. _Nature_ 504, 268 (2013). Article ADS CAS PubMed Google Scholar * Codron, F. Ekman heat transport for slab oceans. _Clim. Dyn._ 38, 379–389
(2012). Article Google Scholar * Lefèvre, F., Lebonnois, S & Forget, F. A three-dimensional photochemical-transport model of the martian atmosphere. _Sixth International Conference on
Mars_. (2003). * Giada, A. et al. Hazy Archean Earth as an Analog for Hazy Earthlike Exoplanets. _American Astronomical Society Meeting Abstracts# 225_, Vol. 225 (2015). * Pavlov, A. A.,
Brown, L. L. & Kasting, J. F. UV shielding of NH3 and O2 by organic hazes in the Archean atmosphere. _J. Geophys. Res. Planets_ 106, 23267–23287 (2001). Article ADS CAS Google Scholar
* Zahnle, K., Claire, M. & Catling., D. The loss of mass‐independent fractionation in sulfur due to a Palaeoproterozoic collapse of atmospheric methane. _Geobiology_ 4, 271–283 (2006).
Article CAS Google Scholar * Claire, M. W. et al. The evolution of solar flux from 0.1 nm to 160 μm: quantitative estimates for planetary studies. _Astrophys. J._ 757, 95 (2012). Article
ADS Google Scholar * Hu, R., Peterson, L. & Wolf, E. T. O2-and CO-rich atmospheres for potentially habitable environments on TRAPPIST-1 planets. _Astrophys. J._ 888, 122 (2020).
Article ADS Google Scholar Download references ACKNOWLEDGEMENTS We thank Daniel Apai, Alex Bixel, Dillon Demo, Zach Grochau-Wright, François Guyot, Betul Kacar, James Kasting, Charles
Lineweaver, Scot Rafkin and Alexander Sotto for discussion; and three anonymous reviewers for their insightful comments and helpful suggestions. This work was supported by Paris Sciences
& Lettres University (IRIS OCAV and PSL-University of Arizona Mobility Program). B.S. acknowledges support from PSL IRIS Origins and conditions for the emergence of life (OCAV) program.
R.F. acknowledges support from the United States National Science Foundation, Dimensions of Biodiversity program (DEB-1831493). AUTHOR INFORMATION Author notes * These authors contributed
equally: Stéphane Mazevet, Régis Ferrière. AUTHORS AND AFFILIATIONS * Institut de Biologie de l’Ecole Normale Supérieure (IBENS), Université Paris Sciences et Lettres, CNRS, INSERM, 75005,
Paris, France Boris Sauterey, Antonin Affholder & Régis Ferrière * International Center for Interdisciplinary Global Environmental Studies (iGLOBES), CNRS, ENS-PSL University, University
of Arizona, Tucson, AZ, 85721, USA Boris Sauterey, Antonin Affholder & Régis Ferrière * Institut de Mécanique Céleste et de Calcul des Ephémérides (IMCCE), Observatoire de Paris,
Université PSL, CNRS, Sorbonne Université, Univ. Lille, F-75014, Paris, France Boris Sauterey, Antonin Affholder & Stéphane Mazevet * LESIA, Observatoire de Paris, Université PSL, CNRS,
Sorbonne Université, Université de Paris, 5 place Jules Janssen, 92195, Meudon, France Benjamin Charnay * Department of Ecology & Evolutionary Biology, University of Arizona, Tucson, AZ,
85721, USA Régis Ferrière Authors * Boris Sauterey View author publications You can also search for this author inPubMed Google Scholar * Benjamin Charnay View author publications You can
also search for this author inPubMed Google Scholar * Antonin Affholder View author publications You can also search for this author inPubMed Google Scholar * Stéphane Mazevet View author
publications You can also search for this author inPubMed Google Scholar * Régis Ferrière View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS
R.F. and S.M. conceived the study. R.F. designed the ecosystem model. A.A. and B.S. refined the model and developed the code. B.C. provided the climate and carbon cycle modules. B.S. and
S.M. designed the Monte-Carlo approach and ran the simulations. B.S. analyzed the results and wrote the first version of the paper. All authors finalized the paper. CORRESPONDING AUTHOR
Correspondence to Boris Sauterey. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PEER REVIEW INFORMATION _Nature Communications_
thanks Janet Siefert and other, anonymous, reviewers for their contributions to the peer review of this work. Peer review reports are available. PUBLISHER’S NOTE Springer Nature remains
neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION PEER REVIEW FILE RIGHTS AND PERMISSIONS
OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or
format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or
other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in
the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Sauterey, B., Charnay, B.,
Affholder, A. _et al._ Co-evolution of primitive methane-cycling ecosystems and early Earth’s atmosphere and climate. _Nat Commun_ 11, 2705 (2020). https://doi.org/10.1038/s41467-020-16374-7
Download citation * Received: 31 October 2019 * Accepted: 28 April 2020 * Published: 01 June 2020 * DOI: https://doi.org/10.1038/s41467-020-16374-7 SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative