Play all audios:
ABSTRACT Aggregation and spreading of α-Synuclein (αSyn) are hallmarks of several neurodegenerative diseases, thus monitoring human αSyn (hαSyn) in animal models or cell cultures is vital
for the field. However, the detection of native hαSyn in such systems is challenging. We show that the nanobody NbSyn87, previously-described to bind hαSyn, also shows cross-reactivity for
the proteasomal subunit Rpn10. As such, when the NbSyn87 is expressed in the absence of hαSyn, it is continuously degraded by the proteasome, while it is stabilized when it binds to hαSyn.
Here, we exploit this feature to design a new Fluorescent Reporter for hαSyn (FluoReSyn) by fusing NbSyn87 to fluorescent proteins, which results in fluorescence signal fluctuations
depending on the presence and amounts of intracellular hαSyn. We characterize this biosensor in cells and tissues to finally reveal the presence of transmittable αSyn in human cerebrospinal
fluid, demonstrating the potential of FluoReSyn for clinical research and diagnostics. SIMILAR CONTENT BEING VIEWED BY OTHERS POTENT INHIBITORS OF TOXIC ALPHA-SYNUCLEIN IDENTIFIED VIA
CELLULAR TIME-RESOLVED FRET BIOSENSORS Article Open access 28 June 2021 SEED AMPLIFICATION ASSAY FOR THE DETECTION OF PATHOLOGIC ALPHA-SYNUCLEIN AGGREGATES IN CEREBROSPINAL FLUID Article 18
January 2023 MONITORING THE INTERACTIONS BETWEEN ALPHA-SYNUCLEIN AND TAU IN VITRO AND IN VIVO USING BIMOLECULAR FLUORESCENCE COMPLEMENTATION Article Open access 22 February 2022 INTRODUCTION
α-synuclein (αSyn) aggregation disorders including Parkinson’s disease (PD), Lewy Body dementia, and multiple system arophy, are a group of disorders characterized by the pathological
occurrence of intracellular inclusions filled with insoluble aggregates of αSyn. These aggregations can be found in various cell types and regions of the central nervous system1. αSyn is a
140 amino-acid long protein that is highly enriched in presynaptic nerve terminals2. Despite extensive efforts, the central molecular and physiological role of αSyn remains to be determined.
Emerging evidence, however, suggests αSyn to be involved in the regulation and possible maturation of synaptic vesicles3, particularly via its role in the assembly of
N-ethylmaleimide-sensitive factor attachment receptor complexes4; key players in synaptic vesicle docking and fusion with the presynaptic membrane. In addition, other functions have been
attributed to αSyn including the regulation of glucose levels5, serving as an antioxidant6 or a chaperone7, and suppressing apoptosis in dopaminergic neurons8. Under normal physiological
conditions, αSyn is found as a monomeric protein existing in equilibrium between an intrinsically disordered form in the cytosol and a membrane bound, α-helical form9. Although still
subjected to debate10, observation of helical αSyn tetramers has also been reported under certain native environments11. It is widely accepted that arrangements of αSyn monomers into
small-to-intermediate oligomeric or larger insoluble assemblies12 is associated with pathogenesis of αSyn aggregation disorders13. Importantly, there is growing evidence for prion-like
cell-to-cell transmission properties of different αSyn conformations where “toxic” αSyn species with seeding properties are suggested to get internalized by a host cell and trigger the
aggregation of endogenous αSyn14,15,16,17,18. There are nevertheless several assumptions, unknown molecular steps and contradictory observations that render the toxic transmission of human
αSyn (hαSyn) an ambiguous notion. These mainly concern pinpointing of the disease underlying αSyn species (e.g., oligomers vs fibrils, phosphorylated αSyn, etc)19, mechanistic steps of the
cell-to-cell transmission paradigm10,20 and the downstream effects of pathological αSyn accumulation that eventually lead to neuronal injury21. Development of tools that can be used for
reliable and reproducible detection and tracking of αSyn in in vitro or in vivo is a very important goal for deciphering molecular mechanisms of disease pathogenesis. A common strategy
employed in studies addressing the cellular uptake, seeding, and transmission phenomena involve manipulation of the αSyn protein itself. Accordingly, αSyn is either pre-labeled with
fluorescent dyes15,22,23, or recombinantly expressed as a fusion protein. Concerning the latter, several different approaches are described which include tagging αSyn with a small epitope
tag (e.g., myc, V5, etc.) for subsequent antibody-mediated immunodetection15,24, expressing it as a fluorescent fusion protein (e.g., YFP-αSyn, DsRed-αSyn, etc.) for direct
visualization25,26 or employing the protein complementation assay principles whereby αSyn is tagged with either N‐terminal or C‐terminal portions of a split fluorescent or bioluminescent
reporter27. Although they have proven to be very helpful in advancing the knowledge about synuclein-related pathologies, one common caveat in such approaches is their limited potential in
recapitulating the natural behavior of untagged native αSyn. For example, it has been shown that fluorescent protein fusions might result in the wrong localization of the studied protein28
or some organic dyes have a propensity to bind to biological membranes29. Considering that a critical role is attributed to the lipid interacting properties of αSyn in its pathological
behavior30, the employment of untagged or native forms of αSyn may be a more-appropriate strategy when attempting to investigate the molecular mechanisms of αSyn pathology. The detection of
untagged, native, or endogenous αSyn in in vitro or in vivo model systems requires the use of other tools. Certain dyes such as Thioflavin S, derived from the histological dye Congo red,
have been commonly utilized for detecting mature protein aggregates in in vivo models of αSyn propagation and PD pathology16,31. However, these dyes have the disadvantage of binding any
protein capable of taking an amyloid conformation and thus do not provide an exclusive labeling of αSyn32. In contrast, antibody-mediated immunodetection is a conventional approach for
specific detection of αSyn. Hereby, a large number of αSyn-targeting antibodies, including conformation specific-ones and engineered antibody fragments are commonly utilized in a variety of
applications33,34. In the recent years, camelid-originated single-domain antibodies, also termed nanobodies, emerged as a promising alternative as they confer several advantages including
recombinant production, enhanced tissue penetration, small size (ideal for super-resolution microscopy), and the ability to be expressed as intrabodies in mammalian cells35,36,37. The latter
feature is particularly attractive as it confers the ability to track and manipulate specific target proteins in living cells38. Nanobodies are increasingly being used for investigation of
diseases associated with protein misfolding and aggregation. Recently, two nanobodies against αSyn have been identified, NbSyn2 (ref. 39) and NbSyn87 (ref. 40), each binding distinct
epitopes at the C-terminal region of αSyn. These have been biochemically well characterized and assessed for their potential therapeutic use by several studies39,40,41,42,43,44. In this
study, we make use of a previously unknown feature of NbSyn87, namely its weak affinity to the 26 S proteasomal subunit protein Rpn10, which is located at the entrance of the proteasome45
and functions as a receptor for poly-ubiquitinated proteins that will undergo proteolysis. We show that this interaction is sufficient to drive a continuous proteasome-mediated degradation
of intracellularly expressed NbSyn87 unless it is bound to hαSyn. The presence of hαSyn, on the other hand, results in the avoidance of the degradation of NbSyn87 by formation of a
stabilized NbSyn87:hαSyn complex. Accordingly, we exploit this mechanism to create and characterize a nanobody-based Fluorescent Reporter for human αSyn (FluoReSyn), which is able to report
the presence or absence of cytosolic hαSyn. Our results demonstrate the unique ability of FluoReSyn to report small amounts of cytosolic hαSyn in cell lines and transduced primary rat
hippocampal neurons. Expression of FluoReSyn in olfactory system of _Xenopus laevis_ also shows its ability to operate and report hαSyn in vivo. Furthermore, cells stably expressing
FluoReSyn (Reporter-cells) report the presence of hαSyn in their cytoplasm after exposing them to human cerebrospinal fluid (CSF) samples. The results presented here indicate that this
biosensor is a valuable instrument for studying the transmission of αSyn and has great potential to be further optimized and validated as a diagnostic tool for αSyn aggregation disorders.
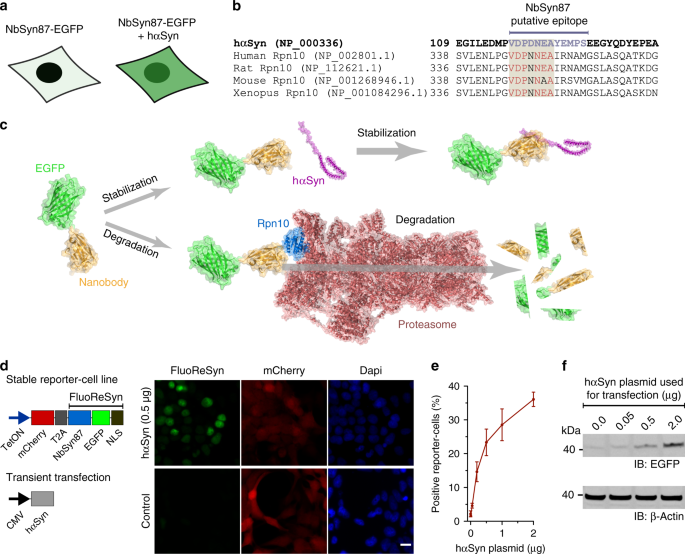
RESULTS REPORTING THE PRESENCE OF UNTAGGED HΑSYN IN THE CYTOPLASM We had previously observed in cells transiently expressing the NbSyn87 (ref. 40) fused to EGFP that their fluorescent signal
correlated with the presence or absence of hαSyn (Fig. 1a). In an attempt to comprehend this observation, we used the Basic Local Alignment Research Tool (BLAST) to find out if the
described hαSyn epitope sequence (VDPDNEAYEMPS)40 that is recognized by the NbSyn87 might be present in another endogenous protein. The BLAST result showed a high % identity (Fig. 1b) to a
subunit of the 26 S proteasome (the 26 S proteasome non-ATPase regulatory subunit four homolog, also known as Rpn10). This protein resides at the entrance of the 26 S proteasome45 and has an
important role in the recognition of poly-ubiquitinated proteins that will be processed in the ubiquitin proteasome-mediated proteolysis (UPP)46. Using a dot-blot assay with purified Rpn10
and hαSyn, we were able to verify that the NbSyn87 can bind weakly to human Rpn10 (Supplementary Fig. 1a, b). Therefore, taking our results together, we hypothesized that the degradation of
NbSyn87 in the absence of hαSyn may be mediated by its weak but continuous recruitment to the proteasome upon binding the endogenous Rpn10 (Fig. 1c). In order to test our hypothesis and
further characterize NbSyn87 in terms of this special feature, we decided to generate a stable cell line expressing NbSyn87 fused to EGFP and have mCherry signal as an expression reporter
using the self-cleavable domain T2A47 (Fig. 1d). In addition, we added a NLS sequence at the C-terminus of NbSyn87-EGFP to concentrate the EGFP signal in the nucleus and gain sensitivity
during imaging. The expression of this protein-chimera was controlled under the tetracycline-inducible promotor system (TetOn). Optimal induction duration to maximize the NbSyn87-EGFP-NLS
expression was determined by analyzing the mCherry reporter signal (Supplementary Fig. 1c). In line with our original observations, we detected a clear nuclear EGFP signal in
doxycycline-induced NbSyn87-EGFP-NLS stable cell line when we transiently transfected them with wild type and untagged hαSyn (Fig. 1d). The strength of this effect was dependent on the
amount of hαSyn present in the cells; detected both by EGFP fluorescence signal intensity analysis (Fig. 1e) and EGFP amounts revealed in western blots (Fig. 1f and Supplementary Fig. 1d).
Doxycycline-induced cells that were not transiently transfected with hαSyn displayed the mCherry reporter signal but no EGFP signal (Fig. 1d). Therefore, we termed the NbSyn87-EGFP-NLS
construct as the Fluorescent Reporter for αSyn (FluoReSyn) and the stably FluoReSyn expressing cell line as the Reporter-cells. PROTEASOME-MEDIATED DEGRADATION OF FLUORESYN In order to
validate our proposed mechanism for the degradation of FluoReSyn, we treated doxycycline-induced Reporter-cells (without expressing hαSyn) over different time periods with the MG132
proteasome inhibitor. MG132 concentration was optimized to provide a strong gain of EGFP and mCherry signal while minimizing its adverse toxic effects on the Reporter-cells (Supplementary
Fig. 2). The results clearly showed that, following treatment with 1 µM of MG132, FluoReSyn started to accumulate in the nuclei of Reporter-cells already from the 4th h on without the
presence of hαSyn (Fig. 2a, b). Induced Reporter-cells untreated with MG132, showed in contrast virtually no FluoReSyn signal throughout the whole duration (Fig. 2a, b). These suggested that
the reporter is regularly produced but also continuously degraded via the proteasome in the cell (fast turnover) under normal conditions. The mCherry and FluoReSyn are produced from a
single mRNA, making a fusion protein that is efficiently cleaved at the T2A domain47. It is expected that this strategy results in stoichiometric amounts of FluoReSyn and mCherry, which
allowed us to normalize the signal of FluoReSyn with the mCherry signal. In induced Reporter-cells untreated with MG132, the relation of FluoReSyn to mCherry signal was maintained during the
16 h experiments (Fig. 2c). However, when using MG132, already after 2 h, the ratio between FluoReSyn signal and mCherry doubled and kept growing over time, showing that the accumulation of
FluoReSyn exceeds that of mCherry and that the former gets particularly enriched in the Reporter-cells when the proteasome machinery is inhibited (Fig. 2c). This observation suggests that
the proteasomal degradation is particularly accelerated for FluoReSyn under normal conditions (uninhibited proteasome machinery), and thus substantiates our proposition that the nanobody
NbSyn87 specifically targets the proteasome and is degraded by it. FLUORESYN REPORTS HΑSYN IN VIVO As Rpn10 is a well-conserved protein across different species (Fig. 1b), we presumed that
our proposed mechanism should also operate in a model organism that lacks endogenous hαSyn and possesses the conserved Rpn10 epitope recognized by NbSyn87. Accordingly, we chose to validate
the proposed mechanism in living _Xenopus laevis_ tadpoles, a time- and cost-efficient model organism48. _Xenopus laevis_ expresses endogenously the same Rpn10 epitope needed for FluoReSyn
to operate and offers a straightforward electroporation-mediated approach for gene delivery to the olfactory receptor neurons in the olfactory epithelium of living animals49 (Fig. 3a, b).
Accordingly, the plasmid encoding for FluoReSyn was electroporated either alone or together with a plasmid encoding for hαSyn fused to mCherry into the right or left nostrils of anesthetized
tadpoles, respectively. By in vivo imaging of tadpoles with two-photon microscopy we observed many GFP-positive nuclei co-localizing with the mCherry signal (hΑSyn) in the left nostrils of
the animals (Fig. 3c, e, g, high magnification example on 3j), which was clearly in contrast with the right nostrils presenting seldom any GFP-positive nucleus (Fig. 3d, f, h). Analysis of
the distribution of GFP fluorescence intensity further confirmed the clear distinction between left and right nostrils (Fig. 3i). Altogether, these data further validated our previous
conclusions by demonstrating that the same mechanism seems to be operational in vivo whereby FluoReSyn is stabilized upon hαSyn binding and cleared from the cell by proteasome-mediated
degradation in the absence of this interaction. THE SPECIFICITY OF THE FLUORESYN FOR HΑSYN As a next step, we assessed whether FluoReSyn binds specifically to human αSyn or has an affinity
towards other synuclein species. For this purpose, Reporter-cells were transiently transfected with plasmids encoding for hαSyn, human ßSyn (hßSyn), or rat αSyn (rαSyn). We analyzed the
proportion of cells with a positive EGFP signal by epifluorescence microscopy (Fig. 4). The expressions of the different synuclein species were controlled by immunostaining with an antibody
that recognizes all transfected variants. Although no significant differences were observed among the different synuclein species in terms of their expression (Supplementary Fig. 3a), only
in hαSyn-transfected Reporter-cells a substantial proportion of the cells were positive for the FluoReSyn signal (Fig. 4b). This result, therefore, proposes that FluoReSyn binds primarily to
human αSyn. It is important to note that the epitope sequence of the hαSyn recognized by NbSny87 has a great identity with the rαSyn sequence with the epitopes differing from each other
only by two residues (Supplementary Fig. 3b). It is noteworthy, that FluoReSyn seems to bind with higher affinity to Rpn10 than to the rαSyn. DETECTION OF HΑSYN UPTAKE FROM THE CULTURE
MEDIUM In relation to the pathological αSyn transmission phenomenon10, we evaluated the ability of FluoReSyn to report the entry of foreign hαSyn into the cellular cytosol. For this purpose,
purified recombinant hαSyn was administered to the culture mediums of induced Reporter-cells either on its own or in a mixed state with a cationic liposome reagent (i.e., RNAiMAX) to
enhance the protein uptake as suggested previously50,51. We observed that, if hαSyn was associated with RNAiMAX, it manages to get into the cytosol and stabilize FluoReSyn as evidenced by
the accumulated EGFP signal in the nucleus (Fig. 5). On the other hand, administrating hßSyn associated with RNAiMAX or just naked hαSyn generated only background levels (i.e., induced
Reporter-cells not exposed to anything) of EGFP-positive cells, which suggests that hαSyn on its own failed to go across the cell plasma membrane efficiently. Subsequently, we setup a
co-culture of Reporter-cells and HEK293 cells stably expressing untagged and wild-type hαSyn under an inducible promotor (TetON-hαSyn cells; Supplementary Fig. 4) to investigate if mammalian
produced hαSyn was able to leave the cells and then enter into the neighboring Reporter-cells. As a control condition, Reporter-cells were co-cultured with wild-type HEK293 cells, which
lack endogenous hαSyn expression. The analysis of co-cultures maintained for 2–5 days revealed no significant differences between the test and control groups failing to confirm the
occurrence of any hαSyn transmission events between the hαSyn producing cells and the Reporter-cells (Supplementary Fig. 5). UPTAKE OF HΑSYN FROM THE CULTURE MEDIUM BY PRIMARY NEURONS As a
next step, we assessed the functionality of FluoReSyn in neurons, by investigating whether it can report the cytosolic presence of hαSyn, after adding recombinant hαSyn to the medium of
primary neuronal cultures. For this purpose, we prepared rat hippocampal neuron cultures and infected them at DIV ~14 with an adeno-associated virus (AAV) encoding for NbSyn87-mCherry-NLS (a
red version of FluoReSyn). We administered both the monomeric and large fibrillar52 forms of hαSyn extracellularly to the culture medium of FluoReSyn-transduced hippocampal neurons. The
analysis suggested that naked monomeric hαSyn can reach the cytosol of neurons and can produce a detectable FluoReSyn signal in their nuclei (Fig. 6a, b). This was in contrast to the large
hαSyn fibrils, which failed to induce FluoReSyn signals above that of the control neurons (Fig. 6b). Based on our previous observations with the Reporter-cell line, where we also
administered the synuclein species pre-mixed with the cationic lipid RNAiMAX (Fig. 6a, c). The results revealed a higher proportion of neurons with FluoReSyn positive nuclei when hαSyn
monomers were pre-mixed with the cationic lipids from RNAiMAX and added to the culture medium (Fig. 6a, c). It was interesting to observe that both, the naked monomers (Fig. 6b) and those
complexed with lipids (Fig. 6c) were able to go across the neuron plasma membranes. This is clearly distinctive to the HEK293-based Reporter-cells, which displayed internalization of hαSyn
only when complexed to cationic lipids (Fig. 5b). The use of cationic lipids did not influence the outcome for fibrils. Similar to the previous observation with uncoated fibrils (Fig. 6b),
no FluoReSyn signal was detected in neurons exposed to RNAiMAX-hαSyn fibril complexes (Fig. 6c). Importantly, we showed that NbSyn87 can clearly bind to our in vitro generated fibrils (Fig.
6d), which also confirmed previous reports40. Thereby, we can exclude that the negative observations were owing to the inability of FluoReSyn to detect our fibrils (Fig. 6d), but are most
likely a proof that fibrils have not succeeded in entering the cytoplasm of neurons. DETECTION OF HΑSYN IN CSF SAMPLES Confident that FluoReSyn can reliably report cytosolic hαSyn, we
decided to evaluate the ability of the Reporter-cells to detect hαSyn species in human-originated biological samples. The main rationale here was to explore the potential usability of this
cellular system for future diagnostic purposes. Therefore, we exposed induced Reporter-cells cultured on a 96-well plate to CSF samples from 42 individuals diagnosed with variable
neurological disorders that were unrelated to αSyn aggregation disorders. Reporter-cells not exposed to CSF, as well as wild-type HEK293 cells exposed and not exposed to CSF, were used as
negative controls. Cells were fixed 24 h post treatment and dozens of images of randomized and non-overlapping locations were automatically acquired per well (Fig. 7a). Data analysis showed
a small but clear trend of positive Reporter-cells that were incubated with human CSF (Fig. 7b). We also correlated the specific clinical diagnosis of each patient and the total αSyn
concentrations in their CSF to the Reporter-cells activity, as displayed in Supplementary Fig. 6. As controls, Reporter-cells not exposed to CSF or wild-type HEK293 cells (not producing any
FluoReSyn chimera) incubated with or without CSF, all displayed background levels of positive signal (Fig. 7b). This observation reassured that the small percentage of positive
Reporter-cells observed upon CSF exposure is a specific and trustworthy response to some forms of hαSyn present in human CSF that can reach the cytosol of our model Reporter-cells.
Altogether, this result not only suggests that human CSF may contain a transmittable form of αSyn that is capable of entering into cells but also opens the possibility to optimize this
system for generating a unique cell-based diagnostic tool for αSyn aggregation disorders. DISCUSSION Here, we present for the first time a unique feature of the NbSyn87, namely, its natural
tendency to bind to the proteasomal subunit Rpn10 that leads to its own degradation and eventual clearance from the cell cytoplasm in the absence of hαSyn. This special feature allowed us to
develop FluoReSyn, the nanobody-based fluorescent reporter for hαSyn, which is capable of detecting the presence or absence of hαSyn in the cellular cytoplasm. Furthermore, our
Reporter-cells stably expressing FluoReSyn were able to detect a transmittable form of hαSyn present in human CSF. Uptake of toxic αSyn species by cells and their subsequent intracellular
trafficking is a crucial part of the proposed αSyn transmission pathology20. Accordingly, we first characterized and assessed the potential of FluoReSyn Reporter-cells to be used as a
research tool for investigating the transmission phenomenon. Our results after introducing recombinant αSyn to the culture medium suggested that the Reporter-cells could reliably report the
uptake of extracellular αSyn. Similar to other reports51,53,54, we did not detect direct cytoplasmic internalization of naked monomeric hαSyn by the Reporter-cells (derived from HEK293
cells). We rather observed that the cytosolic uptake required assistance with lipid-based elements as also had been shown in other studies51,54. Different from the HEK-based Reporter-cells,
we observed that entry of naked recombinant αSyn monomers into primary neurons expressing FluoReSyn is more likely to occur, and lipid-based facilitating agents like the RNAiMAX make the
entry process even more efficient. These observations expose the influence of the cellular context on the translocation of αSyn from the extracellular to the intracellular space (e.g.,
requirement of distinct receptor interactions or membrane translocators for αSyn)20 and can be interpreted as a reflection of the neuronal nature of αSyn transmission pathology. In line with
this interpretation, we did not observe a transmission event when HEK293-based cells stably transfected with hαSyn were co-cultured with the Reporter-cells. On the other hand, these
transmission events might be highly dependent on the concentration of hαSyn and duration of exposure. With our setup we could not employ higher concentrations of hαSyn in order to avoid
toxic effects to the cells and furthermore, we were limited in terms of exposure durations as a longer maintenance of dividing cells in culture became difficult after several days. In the
current study, we did not observe a positive FluoReSyn signal in transduced neurons following their exposure to fibrillary αSyn. This was not an unexpected observation since fibrils are
large structures (in the µm range) that should not enter the cellular cytosol easily, especially as they are known to not form pore-like structures on membranes55. Accordingly, it is
probable that our observations merely reflected the inability of these bulky structures to penetrate into the neuronal cytosol. It has been proposed that the uptake of larger arrangements
such as oligomers, fibrils, or aggregated αSyn might be mediated by regular endocytosis20,22. In this case, FluoReSyn would not detect compartmentalized αSyn in endocytosed vesicles unless
αSyn finds a way to escape into the cytosol. It has recently been shown that αSyn pre-formed fibrils (pffs; <50 nm of length56) that were internalized by cultured primary neurons remained
confined to endo-lysosomal compartments up to 7 days, without a major escape from the endocytic pathway57. Thus, even small fibrils have difficulties penetrating into the neuronal cytosol.
Finally, these results do not exclude the possibility that other types of large assemblies or fibrils do penetrate into the cell cytosol, and would therefore be detected by the sensor. This
issue could be tested more thoroughly in the future. It is known that many types of fibril-like assemblies with different morphologies and structures can be obtained by changing the
preparation and incubation conditions (e.g., pH, salinity, temperature, presence of modulators)58,59. Structural variabilities of recombinant fibrils were also shown to propagate to in vitro
and in vivo functional properties of the formed assemblies19,58,59. For example, it has been shown that upon reducing the incubation pH, the morphology of formed high-molecular weight αSyn
assemblies shifted from fibrillary to more amorphous with the latter showing reduced in vitro seeding efficiency58. Another study demonstrated that simply by changing from physiological salt
concentrations to a lower salt condition, or by adding a chelating agent to the incubation buffer the appearance of the formed high-molecular weight assemblies of hαSyn changed from
cylindrical to flat. These assemblies, referred to as fibrils or ribbons, respectively, were shown not only to differ in other structural aspects but also in terms of their in vitro-, in
vivo seeding, and propagation properties as well as the degrees of cellular toxicity they induced59. Our results demonstrated that the FluoReSyn works both in a cell line and primary
neurons, and can report the uptake of recombinant αSyn monomers. Introducing recombinant or tissue derived αSyn extracellularly to in vitro cell cultures is a common approach for
investigating the internalization and subsequent seeding activities of αSyn in the context of transmission paradigm23,60,61. However, the readily available cellular models and tools are
generally limited in terms of offering an unambiguous and spatially resolved discernment of internalized αSyn from the extracellularly applied αSyn seeds57. Therefore, there is a necessity
to develop novel tools that would enable selective visualization of internalized αSyn and deliver a quantitative characterization of αSyn uptake phenomenon57. In a recent study where
neuronal cultures were exposed to GFP-tagged αSyn pre-formed fibrils (pffs), a membrane-impermeable fluorescence quencher dye was used to exclusively quench the fluorescence of extracellular
fibrils, thus enabling the selective imaging of only internalized seeds57. Similarly, we propose that, FluoReSyn is an optimal tool for this purpose as it can exclusively detect cytosolic
αSyn. A major advantage of our system is that it avoids the necessity of employing tagged or covalently conjugated forms of αSyn, which as discussed previously may not recapitulate the
normal behavior of native, untagged αSyn. Accordingly, provided that the nanobody epitope is exposed, FluoReSyn can be used for tracking the behavior of any endogenous species of native αSyn
obtained from human materials. Our data suggest that FluoReSyn is a useful research tool not only for neuronal cultures, but also for in vivo set-ups like in our experiments with living
_X. Laevis_. Besides, the FluoReSyn’s specificity to hαSyn would be of advantage when studying hαSyn transmission in animal models (e.g., mice, rat) as any interference from endogenous αSyn
present in rodents can be disregarded. As a next step in further characterizing the cellular reporter system, we evaluated its ability to detect human-originated αSyn with the future
perspective of developing it further into a diagnostic tool62,63,64,65. Here, we used CSF samples from a readily available cohort of individuals (42). The results showed that our
Reporter-cells could indeed detect specific species of αSyn in human CSF, which are able to enter into their cytoplasm. Considering our observations with recombinant αSyn monomers that
required a lipid coating agent for entering the Reporter-cells, it is plausible that the αSyn molecules we detected in CSF samples were in vesicular structures such as exosomes as already
suggested by others66,67, or were species other than monomers, such as oligomers with different proposed means of entering the cells68,69,70,71. The presence of αSyn in CSF can be measured
biochemically (e.g., ELISA)72, and many efforts have been directed at developing assays for detection of CSF-originated αSyn as a biomarker73,74. It is important to emphasize that our system
provides extra information by detecting hαSyn forms that are able to get into the cytosol of cells. Therefore, it is plausible that we are detecting the species that are more prone to
transmission related pathology. This is particularly relevant in the light of emerging evidence, which suggests that particular species of αSyn (e.g., oligomers, aggregates) that are
associated with toxicity can serve as far-better biomarkers than total αSyn in CSF74. Although our initial results in Fig. 1 show that the FluoReSyn signal correlates to the amount of αSyn,
our Reporter-cells are not yet quantitative enough to precisely determine the concentrations of transmittable αSyn in human CSF. Nevertheless, this is a fascinating first proof-of-concept.
To the best of our knowledge, our cellular hαSyn reporter system is so far the sole approach developed for the detection of transmittable hαSyn species present in human body fluids. We
believe that major optimizations can be performed to this system to increase its sensitivity and accuracy. These enhancements might include employment of a faster maturing and brighter EGFP
variant, tuning of the FluoReSyn sensitivity biochemically, and destabilizing the antigen-unbound NbSyn87 (ref. 75). Once these optimizations are performed and validated, a logical next step
would be the generation of a knock-in (KI) mice, to thus generate an animal model for studying the mechanical and molecular aspects of αSyn transmission. Using primary cells from these KI
mice, it will then be possible to establish a neuron-based reporter system, which would presumably hold more promise in a diagnostic context. We hope such efforts might help to achieve more
sensitive and quantitative read-outs, which would in turn pave the way for the development of an accurate and reliable diagnostic or prognostic tool for αSyn associated disorders. METHODS
PLASMID TRANSFECTIONS AND VIRUS INFECTIONS Transfections with pcDNA 3.1 (+) vectors (Thermo Fisher Scientific) encoding for hαSyn (accession number NM_000345.4), human β-synuclein (hβSyn;
accession number NM_001001502.3) or rat αSyn (rαSyn; accession number NM_019169.2) sequences were performed with Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific) and Opti-MEM I
Reduced Serum Medium (Opti-MEM from Gibco, Thermo Fisher Scientific, Waltham, MA, USA) according to manufacturer’s instructions. Transfected cells were typically used after 48 h. The AAV
coding for NbSyn87 fused to mCherry and nuclear localization signal (NLS) sequences (NbSyn87-mCherry-NLS) was kindly provided by Dr. Sebastian Kügler, Department of Neurology; Viral Vectors
Laboratory, University Medical Center Göttingen, Germany. PROTEIN PURIFICATION Rpn10, hαSyn, hβSyn were produced using NEB Express Competent _Escherichia coli_ (New England BioLabs Inc.,
Ipswich, MA, USA). Bacteria were grown overnight with the plasmid of interest in Lysogeny Broth (Sigma-Aldrich) and the respective antibiotics. Next day, cells were further cultured in
Terrific Broth (Sigma-Aldrich) and antibiotics until OD was ~2–3 at 37 °C and induced with 0.8 mM IPTG for 4 h. After adding 5 mM EDTA, cells were harvested by centrifugation at ~ 3000 × _g_
for 30 min at 10 °C and frozen at −20 °C until further processing. Pellets were resuspended on ice with 1 mM DTT, 25 mM Imidazole, and 1 mM PMSF in a His-binding buffer (50 mM HEPES pH 8.0,
500 mM NaCl, 5 mM MgCl2 and 10% glycerol) and bacteria were lysed by sonication on ice. Cell debris was separated by centrifugation at ~ 15,000 × _g_ for 1 h at 4 °C. Clear supernatant was
incubated with 2 ml of pre-equilibrated Ni-beads (cOmplete His-Tag Purification Resin, Roche, Switzerland) for 1 h and then transferred to a sigma column to be washed consecutively with
His-binding buffer (50 mM HEPES pH 8.0, 1000 mM NaCl, 10 mM MgCl2, 25 mM Imidazole and 5% glycerol) and low-Salt-pH buffer (50 mM HEPES pH 7.5, 500 mM NaCl, 5 mM MgCl2, 25 mM Imidazole, 5%
glycerol) with each of them containing 25 mM Imidazole. Finally, protein was eluted by 500 mM Imidazole and His-Tag was cleaved with SUMO protease and removed by reverse binding to nickel
beads. Pure proteins were desalted into PBS, purity was confirmed by polyacrylamide gels (PAGE) and concentrations were determined using nanodrop spectrophotometer considering the protein
molecular weight and extinction coefficient. NBSYN87 COUPLING TO ALEXA647 FLUOROPHORE NbSyn87 protein was obtained from a custom production service offered by NanoTag Biotechnologies GmbH
(Göttingen, Germany), and the nanobody was equipped with one ectopic cysteine at its C-terminal. 1 mg of pure NbSyn87 was reduced by adding TCEP (Sigma-Aldrich) to a final concentration of 5
mM for 1 h. The reduced sample was then desalted using gravity columns Nap10 (GE Healthcare Life Sciences) in nitrogen bubbled PBS (pH 7.4) and immediately added to five molar excess of
maleimide-functionalized Alexa647 (Thermo Scientific) for 1 h. Excess of free dye was separated from the conjugated nanobody with an Äkta HPLC equipped with a Superdex 75 increase column (GE
Healthcare Life Sciences). GENERATION OF HΑSYN FIBRILS Monomeric hαSyn was expressed recombinantly in _E. coli_ BL21 (DE3) and purified by anion exchange and size-exclusion
chromatography58. Cell lysis was conducted by French press (Avestin EmulsiFlex-C3) with lysis buffer (10 mM Tris-HCl, pH 8, 1 mM EDTA, 1 mM PMSF), 20 mL per 1 L of cell culture. The lysate
was then heated up to 96 °C for 30 min in a water bath and centrifuged afterwards for 30 min at 4 °C with 22,000 × _g_ (Beckman Coulter, JA 25-5 rotor). Streptomycin was added at a final
concentration of 10 mg/mL and incubated for 15 min. Following another centrifugation step, the soluble protein was precipitated by ammonium sulfate for 15 min at 4 °C. Protein pellet was
dialyzed overnight against 25 mM Tris-HCl, pH 7.7 and loaded on an anion exchange column (GE Healthcare, Mono Q 5/50 GL). Protein was eluted with 300 mM NaCl. Monomeric hαSyn purity and was
achieved by size-exclusion chromatography (GE Healthcare, Superdex 75 10/300 GL) using 50 mM HEPES, pH 7.4, 100 mM NaCl, and 0.02% NaN3. The protein was sterile filtered (0.22-μm) and stored
at 1 mM at −80 °C. Monomeric hαSyn in 50 mM HEPES, pH 7.4, 100 mM NaCl, and 0.02% NaN3 was centrifuged at 84,000 × _g_ for 1 h at 4 °C. The supernatant was filtrated through 0.22 µm
ULTRAFREE-MC centrifugal filter units (Millipore) and adjusted to 0.25 mM protein concentration. Aggregation was performed for 10 days at 37 °C with constant stirring at 200 rpm. Progress of
fibril formation was monitored with a Thioflavin T fluorescence assay76. The fibrils were finally collected by ultracentrifugation at 20 °C, washed twice with 50 mM HEPES, pH 7.4, 100 mM
NaCl and quantified by subtracting the amount of monomeric hαSyn in the supernatant from the total protein used for aggregation. Prior to their use, fibrils were resuspended in the washing
buffer at 0.1 mM protein concentration. PROTEASOME INHIBITION MG132 (Sigma-Aldrich, St. Louis, MO, USA) was administrated at different concentrations (Supplementary Fig. 2) or at 1 μM to the
culture medium for different time intervals (Fig. 2). CSF SAMPLES Study participants consisted of individuals who were in treatment at the Paracelsus Elena Klinik, Kassel, Germany, and had
been diagnosed with a variety of neurological disorders non-related to αSyn aggregation disorders. The study cohort consisted of 23 females and 19 male individuals with a mean age of 70.95 ±
1.51. For a detailed presentation of demographic and clinical features of participants please see Supplementary Table 1. CSF samples from all individuals were collected after the informed
consent of the participant at the Paracelsus Elena Klinik in accordance with the principles of Declaration of Helsinki and following identical standard operating procedures. In brief, CSF
was collected by lumbar puncture in the morning with fasting patients in a sitting position. The samples were centrifuged at 2000 × _g_ for 10 min at room temperature (RT). The
concentrations of αSyn in CSF samples were measured with a validated sandwich ELISA system (mSA1/Syn1-BB; 384 well plate format)72. Aliquots of the supernatants were frozen within 20–30 min
and stored at −80 °C until their use. A total cell count was established in tube 1 (2 mL). Samples with erythrocyte counts >50 cells per μL CSF in tube 1 were excluded from all analyses.
The use of the CSF samples in this study was approved by the ethical committee of the Medical Center Göttingen with the approval numbers 36/7/02 and 9/7/04. CELL LINES Wild-type HEK293 were
grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS, 4 mM l-glutamine and 600 U/ml penicillin-streptomycin (Lonza). Reporter-cells (HEK293 stably
expressing TetON-NbSyn87-EGFP-P2A-mCherry) and hαSyn cells (HEK293 stably expressing TetON-hαSyn) were grown in DMEM supplemented with 10% FBS, 2 mM l-glutamine supplemented with 0.5 µg/ml
puromycin (InvivoGen, San Diego, CA, USA). Both, Reporter-cells and hαSyn cells were produced by Sirion Biotech GmbH (Martinsried, Germany). TetON Induction was performed using 0.5 µg/ml
doxycycline (Sigma-Aldrich, St. Louis, MO, USA) at least 12 h before their use. PRIMARY HIPPOCAMPAL NEURONS Postnatal (P1–P2) pups from Wistar rats were decapitated and the brains were
extracted. The hippocampi were isolated, washed in Hank’s balanced salt solution (HBSS; Invitrogen, Darmstadt, Germany) and incubated in enzymatic digestion solution for 1 h at room
temperature. After washing in HBSS the hippocampi were incubated in inactivation solution for 15 min. After another washing step in Neurobasal A, neurons were mechanically dissociated by
pipetting. In all, 15,000 neurons per well were added to the plating medium (MEM, 10% horse serum, 3.3 mM glucose, 2 mM glutamine) in PLL-coated 96-multiple glass bottom well plates
(SensoPlate, Greiner Bio-One International GmbH, Kremsmünster, Austria) and kept at 37 °C, 5% CO2. After ~ 1 h, when the neurons adhered to the glass bottom, the plating medium was exchanged
with 100 μL Neurobasal A and plates were further cultured at 37 °C, 5% CO2. To maintain healthy cultures, 50 μL of medium was removed every second day and replaced with 50 μL of fresh
Neurobasal A. The primary rat hippocampal neuron cultures were prepared with minor modifications from the original protocol77. Five days after plating, neurons were infected with an AAV
(88e5 TU AAV/5 μl) containing the sequence for NbSyn87 fused to mCherry and NLS sequences (NbSyn87-mCherry-NLS). CELLULAR UPTAKE OF RECOMBINANT HΑSYN FROM THE CULTURE MEDIUM Purified
synuclein proteins were introduced to the medium of (i) neurons cultured on 96-well plates 6 days post infection with AAV NbSyn87-mCherry-NLS and (ii) Reporter-cells cultured on 24-well
plates 48 h (~70,000 cells per well) post-seeding. hαSyn (monomers or fibrils) and hβSyn proteins were diluted in Opti-MEM and incubated with Lipofectamine RNAimax (Invitrogen, Thermo Fisher
Scientific, 2 and 0.3 µl per well for Reporter-cells and neurons, respectively) for 20 min and then administered to the culture medium at a final concentration of 20 µM protein per well.
Cells were fixed 14 h later for imaging and evaluation. DETECTION OF HΑSYN SPECIES IN CSF SAMPLES Wild-type HEK293 and Reporter-cells were seeded (~ 14,000 per well) and induced in a
PLL-coated 96-well plate with glass bottom (SensoPlate, Greiner Bio-One International GmbH, Kremsmünster, Austria). After 24 h of induction, the cells were exposed to CSF samples. In brief,
the medium was partially eliminated leaving 25 µl per well, and supplemented with 50 µl of CSF or culture medium as negative control. Cells were further incubated for 24 h and then fixed by
adding 4% paraformaldehyde in PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4; pH 7.4) overnight at 4 °C, aldehyde groups were quenched with 0.1 M NH4Cl for 15 min and images were
acquired. IMMUNOSTAINING Cells were briefly washed with DPBS (Reporter-cells) or Tyrode buffer (primary rat hippocampal neurons), fixed with 4% paraformaldehyde in PBS for 30 min at RT.
Remaining reactive aldehyde groups were quenched in PBS supplemented with 0.1 M glycine and 0.1 M NH4Cl for 15 min at RT. Cells were permeabilized and unspecific protein binding sites were
blocked with a blocking/permeabilization solution (0.1% Triton X-100 and 2% bovine serum albumin, BSA in PBS) at RT for 15 min. Reporter-cells were incubated with a rabbit polyclonal
anti-α/β-Synuclein antibody (dilution 1:500; Cat. No. 128002, SySy, Göttingen, Germany) and neurons with a polyclonal guinea pig anti-synaptophysin antibody (dilution 1:100 Cat. No. 101004,
SySy, Göttingen, Germany) for 1 h. After three thorough washing steps, cells were incubated with the secondary antibodies (donkey anti-rabbit, dilution 1:500; Cat. No. 711-175-152, Dianova,
Hamburg, Germany or donkey anti-guinea pig IgG labeled with ATTO 647 N, dilution 1:500; Cat. No. N0602-At647N-S, SySy, Göttingen, Germany) for 45 min or 1 h, respectively. The samples were
again subjected to three thorough washing steps with PBS and high-salt PBS (500 mM NaCl, 2.7 mM KCl, 10 mM Na2PO4, 2 mM KH2PO4; pH 7.3–7.4). Before mounting the coverslips or before imaging,
cells were stained with Höchst 33342 (1 µg/ml; Thermo Fisher Scientific). Coverslips were finally mounted in Mowiol mounting media (6 g glycerol, 6 ml deionized water, 12 ml 0.2 M Tris
buffer pH 8.5, 2.4 g Mowiol 4-88 from Merck). In all, 96-well plates were imaged in PBS. PLASMID ELECTROPORATION OF _XENOPUS LAEVIS_ TADPOLES All procedures for animal handling were approved
by the governmental animal care and use office (Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, Oldenburg, Germany, Az.12/0779) and were in accordance with the
German Animal Welfare Act as well as with the guidelines of the Göttingen University Committee for Ethics in Animal Experimentation. _X. laevis_ tadpoles (albinos, stage 53)78 were used for
the in vivo experiments. Injection micropipettes were pulled from borosilicate microcapillaries (Warner instruments; outer diameter: 1.0 mm, inner diameter: 0.58 mm, length 100 mm) using a
horizontal puller (P 1000, Sutter Instruments). Micropipette tips were sharpened at an angle of 20–30° until the pipette tip had a syringe-like shape (Micropipette Beveler 48000; World
Precision Instruments). Micropipettes were filled with 3 µl of plasmid solution/s (600 ng/µl). Cascade blue dextran (3 mM, 10%, Thermo Fisher) was added to the plasmid solution before to be
able to observe dye extrusion under fluorescent illumination. Albino tadpoles were anaesthetized in 0.02% MS-222 (ethyl 3-aminobenzoate methanesulfonate; Sigma-Aldrich; pH: 7.6) for 5 min
until complete immobility and irresponsiveness. Subsequently, the animal was transferred to a moistened dish under a stereomicroscope with brightfield and fluorescent illumination (Olympus
SZX16; light source: X-Cite Series 120 Q, Lumen dynamics). The glass pipettes filled with plasmid solution/s were mounted to a micromanipulator connected to a FemtoJet injection system
(Eppendorf). The micropipette was carefully penetrated into the olfactory mucosa at three to five different locations without injuring major arteries. Up to five pressure pulses of 250–1000
hPa (1 s each) were applied per site. Once homogenous blue fluorescent signal was visible throughout the olfactory mucosa, an external electric field was applied to the olfactory mucosa
using an electroporation setup79. One of the 0.2 mm platinum wire electrodes was positioned in the water-filled nostril, the other one in contact to the skin, laterally to the olfactory
nerve. Trains of three square pulses (20 V, 500 ms duration, and 25 ms delay) were applied four times in alternating polarity (ELP-01D, NPI Electronics; additional capacitor connected in
parallel: Domoport, 3 µF). The entire procedure was performed in >5 min to grant proper anesthesia and avoid dissipation of plasmid solution before electroporation. Following
electroporation, animals were put into water until they woke from anesthesia. After assessment of normal swimming the larvae were left in their aquaria to recover for at least 24 h.
Individual nostrils were electroporated sequentially, with a 1 h recovery period in between. We injected and electroporated two plasmids in the left nostril: one expressing FluoReSyn and the
other hαSyn, both under the CMV promotor. In the right nostril, only the FluoReSyn expressing plasmid was electroporated. IMAGING Conventional epifluorescence images of the Reporter-cells
were obtained with an Olympus IX71 microscope equipped with a 0.5 NA/×20 dry UPlanFL N objective and captured with an Olympus F-View II CCD camera (Olympus, Hamburg, Germany). Experiments
with CSF samples and neurons on 96-well plates were acquired using a Biotek Cytation 3 Imaging Reader (BioTek Instruments, Winooski, VT, USA) equipped with a ×20, Plan Fluorite WD 6.6 NA
0.45 objective, a 465 nm LED cube (Cat# 1225001), EGFP Filter cube (Cat# 1225101), 523 nm LED cube (Cat# 1225003), RFP filter cube (Cat# 1225103) and a 16-bit monochromatic CCD camera (pixel
size 6.45 µm × 6.45 µm). IN VIVO MULTIPHOTON IMAGING OF THE _XENOPUS_ OLFACTORY SYSTEM For in vivo imaging, we anesthetized the electroporated tadpoles in 0.02% MS-222 (ethyl
3-aminobenzoate methanesulfonate; Sigma-Aldrich; pH: 7.6) for 5 min until complete irresponsiveness. The animals were placed into a recess of a silicone-filled recording chamber. The chamber
was filled with water and the animal was mechanically fixed using parafilm. A small window was cut into the parafilm to expose the olfactory organs. Both nostrils were imaged under a
two-photon microscope (Nikon A1R MP) at an excitation wavelength of 920 nm. All animals (_n_ = 9) and nostrils were imaged as 3D image stacks under the same gain and laser settings, to
compare fluorescence intensities of the FluoReSyn. The imaging procedure did not last longer than 10 min and animals were transferred to a big water-filled beaker until they recovered from
anesthesia. CELL LYSATE PREPARATION Reporter-cells were washed briefly with ice-cold DPBS and lysed with 50 µl of lysis buffer per well (50 mM Tris/HCl pH 7.5, 150 mM NaCl, 2 mM EDTA, 0.5%
IgePAL, 0.5% Sodium deoxycholate and freshly added 250 µM PMSF, 10 ng Leupeptin, 10 ng Aprotinin, 1 ng Pepstatin A, 10 ng DNase und 1 µl Halt Protease Inhibitor Cocktail; Thermo Fisher
Scientific). Cell lysates were collected into a pre-cooled tube and were centrifuged for at least 1 h at ~15,000 × _g_ at 4 °C. The supernatant was collected into two tubes, snap-frozen with
liquid nitrogen and stored at −80 °C until needed. WESTERN BLOTTING Reporter-cell lysates were thawed on ice and diluted accordingly to their total protein (determined using BCA assay
(Merk)) to load the same total protein concentration in each lane. Samples were mixed with pre-heated 5× Laemmli buffer (50 mM Tris-HCl, 4% sodium doedecyl sulfate (SDS), 0.01% Serva Blue G,
12% glycerol, pH 6.8, 50 mM DTT) to be further boiled at 95 °C for 10 min, centrifuged and then loaded into previously casted 10–12% PAGE. After the SDS-PAGE run was completed, proteins
were transferred to a nitrocellulose membrane in wet trans blot cell (Biorad) with 400 mA for 2 h at 4 °C while stirring the transfer buffer (25 mM Tris, 192 mM glycine, pH 8.3 and 20%
methanol, and 0.04% SDS). The membrane was incubated for 1 h in blocking buffer (5% Nonfat Dried Milk, 0.1% Tween20 in PBS) and then was further incubated with a mouse monoclonal anti-EGFP
antibody (1:500; Cat. No. A11120, Invitrogen, Thermo Fisher Scientific) or a rabbit polyclonal anti-β-Actin-Cy5 antibody (1:1000; Cat. No. 251003, SySy, Göttingen, Germany). Primary antibody
incubations were performed overnight at 4 °C with constant shaking. The following day, the membrane was washed thoroughly in blocking buffer and incubated with the fluorescently labeled
secondary donkey polyclonal anti-mouse antibody (1:1000; Cat. No. 715-175-150, Dianova) for 1 h at RT. For experiments with the directly labeled anti-β-Actin antibody, the second 1 h
incubation step was omitted. Membrane was washed several times each in 0.1% Tween20 in PBS and imaged using an Amersham Imager 600 (GE Healthcare Life Sciences, Little Chalfont, UK).
DOT-BLOT Proteins were serial diluted in washing buffer (0.05% Tween20 in PBS) and spotted on a nitrocellulose membrane. After the membrane was dried, it was blocked with 2% FBS or BSA, 5%
Nonfat Dried Milk, 0.05% Tween20 in PBS for 1 h under agitation. This was followed by incubation with the fluorescently labeled nanobody NbSyn87-Alexa647 for 1 h. Unbound nanobodies were
washed away by several thorough washing steps with 0.05% Tween20 in PBS for a total duration of 1 h. Finally, images of the membranes were taken with an Amersham Imager 600 (GE Healthcare
Life Sciences, Little Chalfont, UK) to detect the NbSyn87-Alexa647 signal. DATA ANALYSIS AND STATISTICS Image analyses of experiments presented in Figs. 1, 2, 4–7, and Supplementary Figures
were performed with custom-written procedures in Matlab (MathWorks Inc., Natick, MA, USA). Cells were identified automatically based on their Höchst 33342 (Figs. 1, 2, 4–6, Supplementary
Fig.) or mCherry (Fig. 7) signals. The average signal intensity within a cell was calculated and corrected for the background intensity by subtracting the background region of interest from
the average signal intensity. Cells were considered as positive if their background-corrected GFP (Figs. 1, 4, 5, Supplementary Fig. 3, 4) or mCherry (Fig. 6) signal intensity (AU) was above
the mean plus 2 standard deviations of control GFP or mCherry signal intensity, respectively. For Fig. 7, cells were considered as positive if background-corrected GFP signal intensity was
above 400 AU. In Fig. 7, signal intensity was normalized to mCherry to exclude differences in FluoReSyn induction. Graph plotting as well as statistical analyses of data presented in Figs.
1, 2, 4–7, and Supplementary Figures were carried out using custom-written procedures in Matlab, Sigma Plot (Systat Software, San Jose, CA, USA) or GraphPad Prism 5.0 (San Diego, CA, USA).
All values are given as mean ± standard error of the mean from at least three independent experiments. Statistical significance was assessed by one-way ANOVA and Tukey’s Post hoc test.
Images showing FluoReSyn signal in vivo (Fig. 3) were created from multi-channel 3D image stacks acquired with the multiphoton microscope. Autofluorescence from melanophores was removed by
subtracting the maximum intensity z-projections of the blue emission channel (where only autofluorescence was visible) from the green emission channel (with FluoReSyn signal only) using Fiji
“Image Calculator” function80. For quantitative analysis of differences in FluoReSyn fluorescence between nostrils, maximum intensity z-projections of the green emission channel were
processed in Fiji to obtain a binary mask of the regions of interest (nuclei with FluoReSyn signal). Therefore, binary images were created using Li’s method of minimum cross-entropy
thresholding81 followed by four rounds of despeckeling in Fiji80. The resulting binary masks were used to measure mean fluorescence intensity values of all regions of interest in the images
with areas between 20 and 150 px (size range of nuclei). Fluorescence mean intensities were pooled for each condition (FluorReSyn + hαSyn vs. only FluoReSyn) from nine (559 nuclei in total)
animals measured under the same conditions. Scatter plots were created using the Seaborn package in Python (python.org; version 0.9; 10.5281/zenodo.1313201). REPORTING SUMMARY Further
information on research design is available in the Nature Research Reporting Summary linked to this article. DATA AVAILABILITY The data that support the finding of this study is readily
available within this paper, its Supplementary file and in the Source Data file. The data set for Figs. 1e, 2b, c, 3i, 4b, 5b, 6b, c, 7b and Supplementary Fig. 1a–c, 3a, b, 5b, and 6 are
provided in the Source Data file. All data sets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. CODE AVAILABILITY
The customized code generated during and/or analyzed during the current study is available from the corresponding author on reasonable request. REFERENCES * Spillantini, M. G. &
Goedert, M. The α-synucleinopathies: Parkinson’s disease, dementia with Lewy bodies, and multiple system atrophy. _Ann. NY Acad. Sci._ 920, 16–27 (2006). Article ADS Google Scholar *
Jakes, R., Spillantini, M. G. & Goedert, M. Identification of two distinct synucleins from human brain. _FEBS Lett._ 345, 27–32 (1994). Article CAS PubMed Google Scholar *
Lautenschläger, J. et al. C-terminal calcium binding of α-synuclein modulates synaptic vesicle interaction. _Nat. Commun._ 9, 712 (2018). Article ADS PubMed PubMed Central CAS Google
Scholar * Burré, J. et al. Synuclein promotes SNARE-complex assembly in vivo and in vitro. _Science_ 329, 1663–1667 (2010). Article ADS PubMed PubMed Central CAS Google Scholar *
Rodriguez-Araujo, G. et al. Alpha-synuclein elicits glucose uptake and utilization in adipocytes through the Gab1/PI3K/Akt transduction pathway. _Cell Mol. Life Sci._ 70, 1123–1133 (2013).
Article CAS PubMed Google Scholar * Hashimoto, M. et al. alpha-Synuclein protects against oxidative stress via inactivation of the c-Jun N-terminal kinase stress-signaling pathway in
neuronal cells. _J. Biol. Chem._ 277, 11465–11472 (2002). Article CAS PubMed Google Scholar * Ahn, M., Kim, S., Kang, M., Ryu, Y. & Kim, T. D. Chaperone-like activities of
alpha-synuclein: alpha-synuclein assists enzyme activities of esterases. _Biochem. Biophys. Res. Commun._ 346, 1142–1149 (2006). Article CAS PubMed Google Scholar * Jin, H. et al.
Synuclein negatively regulates protein kinase Cδ expression to suppress apoptosis in dopaminergic neurons by reducing p300 histone acetyltransferase activity. _J. Neurosci._ 31, 2035–2051
(2011). Article CAS PubMed PubMed Central Google Scholar * Burré, J., Sharma, M. & Südhof, T. C. α-Synuclein assembles into higher-order multimers upon membrane binding to promote
SNARE complex formation. _Proc. Natl Acad. Sci._ _USA_ 111, E4274–E4283 (2014). Article ADS PubMed CAS PubMed Central Google Scholar * Karpowicz, R. J., Trojanowski, J. Q. & Lee,
V. M.-Y. Transmission of α-synuclein seeds in neurodegenerative disease: recent developments. _Lab. Invest._ 99, 971–981 (2019). Article PubMed PubMed Central Google Scholar * Bartels,
T., Choi, J. G. & Selkoe, D. J. α-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation. _Nature_ 477, 107–110 (2011). Article ADS CAS PubMed
PubMed Central Google Scholar * Fink, A. L. The aggregation and fibrillation of α-synuclein. _Acc. Chem. Res._ 39, 628–634 (2006). Article CAS PubMed Google Scholar * Opazo, F., Krenz,
A., Heermann, S., Schulz, J. B. & Falkenburger, B. H. Accumulation and clearance of α-synuclein aggregates demonstrated by time-lapse imaging. _J. Neurochem_ 106, 529–540 (2008).
Article CAS PubMed Google Scholar * Li, J.-Y. et al. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest host-to-graft disease propagation. _Nat. Med._ 14,
501–503 (2008). Article CAS PubMed Google Scholar * Desplats, P. et al. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. _Proc. Natl
Acad. Sci. USA_ 106, 13010–13015 (2009). Article ADS CAS PubMed PubMed Central Google Scholar * Luk, K. C. et al. Pathological α-synuclein transmission initiates Parkinson-like
neurodegeneration in nontransgenic mice. _Science_ 338, 949–953 (2012). Article ADS CAS PubMed PubMed Central Google Scholar * Masuda-Suzukake, M. et al. Prion-like spreading of
pathological α-synuclein in brain. _Brain_ 136, 1128–1138 (2013). Article PubMed PubMed Central Google Scholar * Brundin, P. & Melki, R. Prying into the prion hypothesis for
Parkinson’s disease. _J. Neurosci._ 37, 9808–9818 (2017). Article CAS PubMed PubMed Central Google Scholar * Peelaerts, W. et al. α-Synuclein strains cause distinct synucleinopathies
after local and systemic administration. _Nature_ 522, 340–344 (2015). Article ADS CAS PubMed Google Scholar * Grozdanov, V. & Danzer, K. M. Release and uptake of pathologic
alpha-synuclein. _Cell Tissue Res._ 373, 175–182 (2018). Article CAS PubMed Google Scholar * Vekrellis, K., Xilouri, M., Emmanouilidou, E., Rideout, H. J. & Stefanis, L. Pathological
roles of α-synuclein in neurological disorders. _Lancet Neurol._ 10, 1015–1025 (2011). Article CAS PubMed Google Scholar * Lee, H. J. et al. Assembly-dependent endocytosis and clearance
of extracellular α-synuclein. _Int. J. Biochem. Cell Biol._ 40, 1835–1849 (2008). Article CAS PubMed Google Scholar * Freundt, E. C. et al. Neuron-to-neuron transmission of α-synuclein
fibrils through axonal transport. _Ann. Neurol._ 72, 517–524 (2012). Article CAS PubMed PubMed Central Google Scholar * Klucken, J., Outeiro, T. F., Nguyen, P., McLean, P. J. &
Hyman, B. T. Detection of novel intracellular α-synuclein oligomeric species by fluorescence lifetime imaging. _FASEB J._ 20, 2050–2057 (2006). Article CAS PubMed Google Scholar *
Hansen, C. et al. α-Synuclein propagates from mouse brain to grafted dopaminergic neurons and seeds aggregation in cultured human cells. _J. Clin. Invest._ 121, 715–725 (2011). Article CAS
PubMed PubMed Central Google Scholar * Hansen, C. et al. A novel α-synuclein-GFP mouse model displays progressive motor impairment, olfactory dysfunction and accumulation of
α-synuclein-GFP. _Neurobiol. Dis._ 56, 145–155 (2013). Article CAS PubMed Google Scholar * Outeiro, T. F. et al. Formation of toxic oligomeric α-synuclein species in living cells. _PLoS
ONE_ 3, e1867 (2008). Article ADS PubMed PubMed Central CAS Google Scholar * Stadler, C. et al. Immunofluorescence and fluorescent-protein tagging show high correlation for protein
localization in mammalian cells. _Nat. Methods_ 10, 315–323 (2013). Article CAS PubMed Google Scholar * Hughes, L. D., Rawle, R. J. & Boxer, S. G. Choose your label wisely:
water-soluble fluorophores often interact with lipid bilayers. _PLoS ONE_ 9, e87649 (2014). Article ADS PubMed PubMed Central CAS Google Scholar * Ugalde, C. L., Lawson, V. A.,
Finkelstein, D. I. & Hill, A. F. The role of lipids in α -synuclein misfolding and neurotoxicity. _J. Biol. Chem._ 294, 9016–9028 (2019). Article PubMed PubMed Central Google Scholar
* Luk, K. C. et al. Intracerebral inoculation of pathological α-synuclein initiates a rapidly progressive neurodegenerative α-synucleinopathy in mice. _J. Exp. Med._ 209, 975–986 (2012).
Article CAS PubMed PubMed Central Google Scholar * Rey, N. L., George, S. & Brundin, P. Spreading the word: Precise animal models and validated methods are vital when evaluating
prion-like behaviour of alpha-synuclein. _Neuropathol. Appl. Neurobiol._ 42, 51–76 (2016). Article CAS PubMed Google Scholar * De Genst, E., Messer, A. & Dobson, C. M. Antibodies and
protein misfolding: from structural research tools to therapeutic strategies. _Biochim. Biophys. Acta_ 1844, 1907–1919 (2014). Article PubMed CAS Google Scholar * Vaikath, N. N. et al.
Antibodies against alpha‐synuclein: tools and therapies. _J. Neurochem._ 150, 612–625 (2019). Article CAS PubMed Google Scholar * Muyldermans, S. Nanobodies: natural single-domain
antibodies. _Annu. Rev. Biochem._ 82, 775–797 (2013). Article CAS PubMed Google Scholar * Maidorn, M., Olichon, A., Rizzoli, S. O. & Opazo, F. Nanobodies reveal an extra-synaptic
population of SNAP-25 and Syntaxin 1A in hippocampal neurons. _mAbs_ 11, 305–321 (2019). Article CAS PubMed Google Scholar * Götzke, H. et al. The ALFA-tag is a highly versatile tool for
nanobody-based bioscience applications. _Nat. Commun._ 10, 4403 (2019). Article ADS PubMed PubMed Central CAS Google Scholar * Caussinus, E., Kanca, O. & Affolter, M. Fluorescent
fusion protein knockout mediated by anti-GFP nanobody. _Nat. Struct. Mol. Biol._ 19, 117–122 (2012). Article CAS Google Scholar * Vuchelen, A. et al. (1)H, (13)C and (15)N assignments of
a camelid nanobody directed against human alpha-synuclein. _Biomol. NMR Assign._ 3, 231–233 (2009). Article CAS PubMed Google Scholar * Guilliams, T. et al. Nanobodies raised against
monomeric α-synuclein distinguish between fibrils at different maturation stages. _J. Mol. Biol._ 425, 2397–2411 (2013). Article CAS PubMed Google Scholar * El-Turk, F. et al. Structural
effects of two camelid nanobodies directed to distinct C-terminal epitopes on α-synuclein. _Biochemistry_ 55, 3116–3122 (2016). Article CAS PubMed Google Scholar * Butler, D. C. et al.
Bifunctional anti-non-amyloid component α-synuclein nanobodies are protective in situ. _PLoS ONE_ 11, e0165964 (2016). Article PubMed PubMed Central CAS Google Scholar * De Genst, E. J.
et al. Structure and properties of a complex of α-synuclein and a single-domain camelid antibody. _J. Mol. Biol._ 402, 326–343 (2010). Article PubMed CAS Google Scholar * Chatterjee, D.
et al. Proteasome-targeted nanobodies alleviate pathology and functional decline in an α-synuclein-based Parkinson’s disease model. _NPJ Park. Dis._ 4, 25 (2018). Article CAS Google
Scholar * Haselbach, D. et al. Long-range allosteric regulation of the human 26S proteasome by 20S proteasome-targeting cancer drugs. _Nat. Commun._ 8, 15578 (2017). Article ADS CAS
PubMed PubMed Central Google Scholar * Hamazaki, J. et al. Rpn10-mediated degradation of ubiquitinated proteins is essential for mouse development. _Mol. Cell Biol._ 27, 6629–6638 (2007).
Article CAS PubMed PubMed Central Google Scholar * Kim, J. H. et al. High cleavage efficiency of a 2A peptide derived from porcine teschovirus-1 in human cell lines, zebrafish and
mice. _PLoS ONE_ 6, e18556 (2011). Article ADS CAS PubMed PubMed Central Google Scholar * Blum, M. & Ott, T. Xenopus: an undervalued model organism to study and model human genetic
disease. _Cells Tissues Organs_ 205, 303–313 (2019). Article CAS Google Scholar * Haas, K., Jensen, K., Sin, W. C., Foa, L. & Cline, H. T. Targeted electroporation in Xenopus
tadpoles in vivo -from single cells to the entire brain. _Differentiation_ 70, 148–154 (2002). Article CAS PubMed Google Scholar * Zuris, J. A. et al. Cationic lipid-mediated delivery of
proteins enables efficient protein-based genome editing in vitro and in vivo. _Nat. Biotechnol._ 33, 73–80 (2015). Article CAS PubMed Google Scholar * Luk, K. C. et al. Exogenous
alpha-synuclein fibrils seed the formation of Lewy body-like intracellular inclusions in cultured cells. _Proc. Natl Acad. Sci. USA_ 106, 20051–20056 (2009). Article ADS CAS PubMed
PubMed Central Google Scholar * Karpinar, D. P. et al. Pre-fibrillar α-synuclein variants with impaired Β-structure increase neurotoxicity in parkinson’s disease models. _EMBO J._ 28,
3256–3268 (2009). Article CAS PubMed PubMed Central Google Scholar * Delenclos, M. et al. Investigation of endocytic pathways for the internalization of exosome-associated oligomeric
alpha-synuclein. _Front. Neurosci._ 11, 172 (2017). Article PubMed PubMed Central Google Scholar * Nonaka, T., Watanabe, S. T., Iwatsubo, T. & Hasegawa, M. Seeded aggregation and
toxicity of α-synuclein and tau: cellular models of neurodegenerative diseases. _J. Biol. Chem._ 285, 34885–34898 (2010). Article CAS PubMed PubMed Central Google Scholar * Kim, H. Y.
et al. Structural properties of pore-forming oligomers of α-synuclein. _J. Am. Chem. Soc._ 131, 17482–17489 (2009). Article CAS PubMed Google Scholar * Volpicelli-Daley, L. A., Luk, K.
C. & Lee, V. M. Y. Addition of exogenous α-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous α-synuclein to Lewy body and Lewy neurite-like
aggregates. _Nat. Protoc._ 9, 2135–2146 (2014). Article CAS PubMed PubMed Central Google Scholar * Karpowicz, R. J. et al. Selective imaging of internalized proteopathic -synuclein
seeds in primary neurons reveals mechanistic insight into transmission of synucleinopathies. _J. Biol. Chem._ 292, 13482–13497 (2017). Article CAS PubMed PubMed Central Google Scholar *
Hoyer, W. et al. Dependence of α-synuclein aggregate morphology on solution conditions. _J. Mol. Biol._ 322, 383–393 (2002). Article CAS PubMed Google Scholar * Bousset, L. et al.
Structural and functional characterization of two alpha-synuclein strains. _Nat. Commun._ 4, 2575 (2013). Article ADS PubMed CAS Google Scholar * Holmes, B. B. et al. Heparan sulfate
proteoglycans mediate internalization and propagation of specific proteopathic seeds. _Proc. Natl Acad. Sci. USA_ 110, E3138–E3147 (2013). Article CAS PubMed PubMed Central Google
Scholar * Brahic, M., Bousset, L., Bieri, G., Melki, R. & Gitler, A. D. Axonal transport and secretion of fibrillar forms of α-synuclein, Aβ42 peptide and HTTExon 1. _Acta Neuropathol._
131, 539–548 (2016). Article CAS PubMed PubMed Central Google Scholar * Rizzo, G. et al. Accuracy of clinical diagnosis of dementia with Lewy bodies: a systematic review and
meta-analysis. _J. Neurol. Neurosurg. Psychiatry_ 89, 358–366 (2018). Article PubMed Google Scholar * Rizzo, G. et al. Accuracy of clinical diagnosis of Parkinson disease: a systematic
review and meta-analysis. _Neurology_ 86, 566–576 (2016). Article PubMed Google Scholar * Jellinger, K. A. Accuracy of clinical diagnosis of Parkinson disease: a systematic review and
meta-analysis. _Neurology_ 87, 237–238 (2016). Article PubMed Google Scholar * Majbour, N. K. et al. Longitudinal changes in CSF alpha-synuclein species reflect Parkinson’s disease
progression. _Mov. Disord._ 31, 1535–1542 (2016). Article CAS PubMed Google Scholar * Stuendl, A. et al. Induction of α-synuclein aggregate formation by CSF exosomes from patients with
Parkinson’s disease and dementia with Lewy bodies. _Brain_ 139, 481–494 (2016). Article PubMed Google Scholar * Shi, M. et al. Plasma exosomal α-synuclein is likely CNS-derived and
increased in Parkinson’s disease. _Acta Neuropathol._ 128, 639–650 (2014). Article CAS PubMed PubMed Central Google Scholar * Tsigelny, I. F. et al. Role of α-synuclein penetration into
the membrane in the mechanisms of oligomer pore formation. _FEBS J._ 279, 1000–1013 (2012). Article CAS PubMed PubMed Central Google Scholar * Volles, M. J. & Lansbury, P. T.
Vesicle permeabilization by protofibrillar α-synuclein is sensitive to Parkinson’s disease-linked mutations and occurs by a pore-like mechanism. _Biochemistry_ 41, 4595–4602 (2002). Article
CAS PubMed Google Scholar * Lashuel, H. A., Hartley, D., Petre, B. M., Walz, T. & Lansbury, P. T. Neurodegenerative disease: amyloid pores from pathogenic mutations. _Nature_ 418,
291 (2002). Article ADS CAS PubMed Google Scholar * Stöckl, M., Claessens, M. M. A. E. & Subramaniam, V. Kinetic measurements give new insights into lipid membrane permeabilization
by α-synuclein oligomers. _Mol. Biosyst._ 8, 338–345 (2012). Article PubMed Google Scholar * Mollenhauer, B. et al. α-Synuclein and tau concentrations in cerebrospinal fluid of patients
presenting with parkinsonism: a cohort study. _Lancet Neurol._ 10, 230–240 (2011). Article CAS PubMed Google Scholar * Maass, F., Schulz, I., Lingor, P., Mollenhauer, B. & Bähr, M.
Cerebrospinal fluid biomarker for Parkinson’s disease: an overview. _Mol. Cell Neurosci._ 97, 60–66 (2019). Article CAS PubMed Google Scholar * Parnetti, L. et al. CSF and blood
biomarkers for Parkinson’s disease. _Lancet Neurol._ 18, 573–586 (2019). Article CAS PubMed Google Scholar * Tang, J. C. Y. et al. Detection and manipulation of live antigen-expressing
cells using conditionally stable nanobodies. _eLife_ 5, e15312 (2016). Article PubMed PubMed Central CAS Google Scholar * Hoyer, W., Cherny, D., Subramaniam, V. & Jovin, T. M.
Impact of the acidic C-terminal region comprising amino acids 109-140 on α-synuclein aggregation in vitro. _Biochemistry_ 43, 16233–16242 (2004). Article CAS PubMed Google Scholar *
Kaech, S. & Banker, G. Culturing hippocampal neurons. _Nat. Protoc._ 1, 2406–2415 (2006). Article CAS PubMed Google Scholar * Nieuwkoop P. & Faber, J. _Normal Table of Xenopus
Laevis (Daudin)_ (Garland Science, 1994). * Weiss, L., Offner, T., Hassenklöver, T. & Manzini, I. Dye electroporation and imaging of calcium signaling in Xenopus nervous system. _Methods
Mol. Biol._ 217–231, 2018 (1865). Google Scholar * Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. _Nat. Methods_ 9, 676–682 (2012). Article CAS PubMed
Google Scholar * Li, C. & Lee, C. Minimum cross entropy thresholding. _Pattern Recognit._ 26, 617–625 (1993). Article Google Scholar Download references ACKNOWLEDGEMENTS F.O.,
F.B.B., I.M., and T.O. were supported by the _Deutsche Forschungsgemeinschaft_ (DFG) through Cluster of Excellence Nanoscale Microscopy and Molecular Physiology of the Brain (CNMPB). We
thank Dr. Sebastian Kügler for providing us the AAVs and helpful discussions. We thank Karin Giller, Melanie Wegstroth, Christina Schäfer, and Nicole Hartelt for excellent technical help.
Supported by the DFG under Germany’s Excellence Strategy - EXC 2067/1- 390729940. Supported in part by the DFG through grant SFB1286/Z3 to S.O.R. AUTHOR INFORMATION Author notes * These
authors contributed equally: Fitnat Buket Basmanav, Felipe Opazo. AUTHORS AND AFFILIATIONS * Department of Neuro- and Sensory Physiology, University Medical Center Göttingen, D-37073,
Göttingen, Germany Christoph Gerdes, Eugenio F. Fornasiero, Nora Wender, Hannes Verbarg, Silvio O. Rizzoli, Fitnat Buket Basmanav & Felipe Opazo * Center for Biostructural Imaging of
Neurodegeneration (BIN), University Medical Center Göttingen, D-37073, Göttingen, Germany Natalia Waal & Felipe Opazo * Institute of Animal Physiology, Department of Animal Physiology
and Molecular Biomedicine, Justus-Liebig University Giessen, 35390, Giessen, Germany Thomas Offner & Ivan Manzini * Institute of Neurophysiology and Cellular Biophysics, University of
Göttingen, Göttingen, Germany Thomas Offner & Ivan Manzini * Department of Neurosurgery, University Medical Center Göttingen, D-37075, Göttingen, Germany Claudia Trenkwalder *
Paracelsus-Elena-Klinik, Klinikstraße 16, 34128, Kassel, Germany Claudia Trenkwalder & Brit Mollenhauer * Department of Neurology, University Medical Center Göttingen, D-37075,
Göttingen, Germany Brit Mollenhauer * German Center for Neurodegenerative Diseases (DZNE), Von-Siebold-Str. 3a, 37075, Göttingen, Germany Timo Strohäker & Markus Zweckstetter *
Department for NMR-based Structural Biology, Max Planck Institute for Biophysical Chemistry, 37077, Göttingen, Germany Markus Zweckstetter & Stefan Becker * Cluster of Excellence
“Multiscale Bioimaging: from Molecular Machines to Networks of Excitable Cells” (MBExC), University of Goettingen, Göttingen, Germany Silvio O. Rizzoli * Campus Laboratory for Advanced
Imaging, Microscopy and Spectroscopy, University of Göttingen, D-37073, Göttingen, Germany Fitnat Buket Basmanav Authors * Christoph Gerdes View author publications You can also search for
this author inPubMed Google Scholar * Natalia Waal View author publications You can also search for this author inPubMed Google Scholar * Thomas Offner View author publications You can also
search for this author inPubMed Google Scholar * Eugenio F. Fornasiero View author publications You can also search for this author inPubMed Google Scholar * Nora Wender View author
publications You can also search for this author inPubMed Google Scholar * Hannes Verbarg View author publications You can also search for this author inPubMed Google Scholar * Ivan Manzini
View author publications You can also search for this author inPubMed Google Scholar * Claudia Trenkwalder View author publications You can also search for this author inPubMed Google
Scholar * Brit Mollenhauer View author publications You can also search for this author inPubMed Google Scholar * Timo Strohäker View author publications You can also search for this author
inPubMed Google Scholar * Markus Zweckstetter View author publications You can also search for this author inPubMed Google Scholar * Stefan Becker View author publications You can also
search for this author inPubMed Google Scholar * Silvio O. Rizzoli View author publications You can also search for this author inPubMed Google Scholar * Fitnat Buket Basmanav View author
publications You can also search for this author inPubMed Google Scholar * Felipe Opazo View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS
C.G., T.O., and E.F.F. designed and conducted experiments, analyzed and interpreted data, and contributed with the writing of the manuscript. N. Waal and H.V. designed and performed
experiments, analyzed and interpreted data. N. Wender initially characterized the sensor, designed the Reporter-cell lines, designed and performed experiments, analyzed and interpreted data.
I.M. contributed to the interpretation of data and scientific discussions. C.T. and B.M. provided the human CSF samples. T.S., M.Z., and S.B. prepared and provided αSyn fibrils. S.O.R.
analyzed and interpreted the data, contributed to the supervision of the study and scientific discussions. F.B.B. designed and performed experiments, analyzed and interpreted the data,
supervised the study and wrote the manuscript. F.O. conceived the project, designed and performed experiments, analyzed and interpreted the data, supervised the study and wrote the
manuscript. CORRESPONDING AUTHOR Correspondence to Felipe Opazo. ETHICS DECLARATIONS COMPETING INTERESTS F.O. and S.O.R. are shareholders of NanoTag Biotechnologies GmbH. All other authors
declare no competing interests. ADDITIONAL INFORMATION PEER REVIEW INFORMATION _Nature Communications_ thanks Alice Chen-Plotkin, Anne Messer and Serge Muyldermans for their contribution to
the peer review of this work. Peer reviewer reports are available PUBLISHER’S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional
affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION PEER REVIEW FILE REPORTING SUMMARY SOURCE DATA SOURCE DATA RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under
a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate
credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article
are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and
your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this
license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Gerdes, C., Waal, N., Offner, T. _et al._ A nanobody-based
fluorescent reporter reveals human α-synuclein in the cell cytosol. _Nat Commun_ 11, 2729 (2020). https://doi.org/10.1038/s41467-020-16575-0 Download citation * Received: 12 November 2019 *
Accepted: 05 May 2020 * Published: 01 June 2020 * DOI: https://doi.org/10.1038/s41467-020-16575-0 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this
content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative