Play all audios:
ABSTRACT Recurrent lung infections are a common cause of morbidity and mortality in people living with HIV and this is exacerbated in smokers even when administered combination
antiretroviral therapy (cART). The incidence of pneumonia is increased with smoking and treatment interruption and is directly dependent on viral load in patients when adjusted for CD4
counts. CFTR dysfunction plays an important role in aberrant airway innate immunity as it is pivotal in regulating mucociliary clearance (MCC) rates and other antibacterial mechanisms of the
airway. In our earlier work, we have demonstrated that bronchial epithelium expresses canonical HIV receptors CD4, CCR5 and CXCR4 and can be infected with HIV. HIV Tat suppresses CFTR mRNA
and function via TGF-β signaling. In the present study, we demonstrate that cigarette smoke (CS) potentiates HIV infection of bronchial epithelial cells by upregulating CD4 and CCR5
expression. HIV and CS individually and additively suppress CFTR biogenesis and function, possibly explaining the increased incidence of lung infections in HIV patients and its exacerbation
in HIV smokers. SIMILAR CONTENT BEING VIEWED BY OTHERS A BASALLY ACTIVE CGAS-STING PATHWAY LIMITS SARS-COV-2 REPLICATION IN A SUBSET OF ACE2 POSITIVE AIRWAY CELL MODELS Article Open access
27 September 2024 THE ESTABLISHMENT OF COPD ORGANOIDS TO STUDY HOST-PATHOGEN INTERACTION REVEALS ENHANCED VIRAL FITNESS OF SARS-COV-2 IN BRONCHI Article Open access 10 December 2022 HIV
INDUCES AIRWAY BASAL PROGENITOR CELLS TO ADOPT AN INFLAMMATORY PHENOTYPE Article Open access 17 February 2021 INTRODUCTION The advent of combination antiretroviral therapy (cART) has led to
a dramatic decline in morbidity and mortality from human immunodeficiency virus (HIV)/AIDS. However, HIV infection is associated with a greater incidence of pulmonary diseases normally
associated with aging and their presentation at younger ages. Bacterial pneumonia is the most common cause of hospitalization and intensive care unit admissions in people living with HIV1.
HIV infection was independently associated with a significantly higher risk for lung infections2. Moreover, the incidence of pneumonia is increased with smoking and is directly dependent on
viral load in patients when adjusted for CD4 counts3,4, suggesting that viral replication also plays an important role in lung infections. In our earlier work, we have demonstrated that
primary normal human bronchial epithelial (NHBE) cells express canonical HIV receptors and can be infected with HIV. HIV infection of NHBE suppresses components of the mucociliary clearance
(MCC) apparatus. Specifically, we observed that, HIV Tat suppresses cystic fibrosis transmembrane conductance regulator (CFTR) biogenesis and function, infection of differentiated bronchial
epithelium disrupts integrity of the epithelial barrier and infection of differentiating NHBE grown at the Air liquid interface (ALI) suppresses ciliogenesis5. CFTR plays a pivotal role in
airway innate immunity specifically in modulating MCC. MCC is a primary innate defense mechanism of airways that protects the host from the noxious effects of airborne pathogens, pollutants
and allergens6. The bacterial spectrum responsible for HIV pneumonias is very similar to that observed in smokers, chronic obstructive pulmonary disease (COPD) patients and in cystic
fibrosis7,8,9,10. Acquired CFTR dysfunction (and MCC dysfunction) plays an important role in the early pathogenesis of these chronic airway diseases in smokers and COPD patients. Both
cigarette smoke (CS) and HIV Tat mediate its suppressive effects on CFTR via a common pathway involving transforming growth factor-beta (TGF-β) signaling5,11. CS and TGF-β has been shown to
upregulate HIV receptors12,13,14 in different cell types. CS has also been shown to directly enhance HIV replication by reactive oxygen species mediated activation of CYP enzymes15,16. Hence
CS can enhance HIV replication thereby increasing HIV Tat levels and in turn leading to a synergistic CFTR suppression. This is significant since 60% of people living with HIV also smoke
tobacco17. We have shown that CS enhances infection of NHBE ALI cultures by R5-tropic virus. We will also show that a combination of CS and HIV infection leads to an additive suppression of
CFTR mRNA and a near complete suppression of CFTR function. The implications of these observations on airway innate immunity will be discussed. RESULTS CS ENHANCES INFECTION OF NHBE ALI
CULTURES BY R5-TROPIC HIV STRAINS BUT NOT BY X4-TROPIC HIV STRAINS In our earlier report, we have shown that differentiated NHBE cells expresses CD4, CXCR4 and CCR5 receptors and can be
infected with X4- or R5-tropic virus5. Given reports of enhanced HIV loads in smokers18,19, we tried to determine if CS increases HIV infection of NHBE cells. Age and lung matched NHBE
cultures redifferentiated at the ALI were exposed to CS and then infected with 5 ng of HIV BaL (R5-tropic) or HIV IIIB strain (X4-tropic) HIV as described by us earlier5. 16 hours
post-infection, cells were washed with phosphate buffer saline (PBS) four times. The fourth wash was collected for p24 analysis and measured as day 0 to confirm that all input virus had been
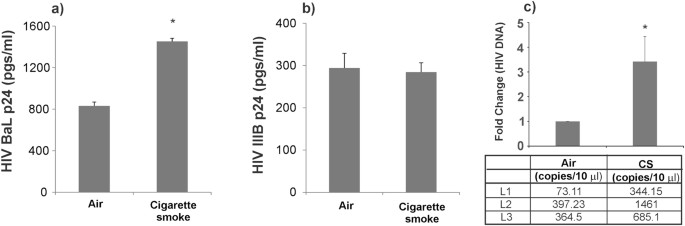
removed. HIV infection of NHBE cells peaks on day 3 and hence basolateral media was collected on day 3 for p24 analysis. As seen in Fig. 1, CS enhances p24 of NHBE cells by the R5-tropic
BaL strain (Fig. 1a) but does not affect infection by the X4-tropic IIIB strain (Fig. 1b). To test if increased p24 expression is due to an increased viral entry, we exposed NHBE ALI
cultures to CS and infected NHBE ALI cultures with the RGH-WT HIV reporter reported by Dahabieh _et al_.20 (and used by us5). RGH virus is an env- single cycle HIV reporter. RGH-WT was
enveloped with R5-tropic env (pBaL.01) as reported by us earlier5. On day 3 post-infection, total DNA was isolated from these cells and HIV DNA was quantitated by qPCR. As seen in Fig. 1c,
increased levels of proviral DNA were observed in NHBE ALI cultures exposed to CS. These data demonstrate that CS enhances viral entry in NHBE ALI cultures. CS UPREGULATES HIV RECEPTORS
Given reports in literature that both, CS and TGF-β (induced by CS) can upregulate HIV receptors12,13,14 in different cell types, we tried to determine if CS also affects expression of these
receptors in NHBE ALI cultures. NHBE cultures redifferentiated at the ALI were exposed to CS and the expression of canonical HIV receptors CD4, CCR5 and CXCR4 were determined by western
blot analysis. As seen in Fig. 2, CS significantly increases expression of the primary HIV receptor CD4 (Fig. 2a). In agreement with reports that have demonstrated that CS enhances CCR5
expression12,13, we also noticed a significant increase in CCR5 expression in NHBE ALI cultures exposed to CS (Fig. 2b). However, CS does not enhance expression of CXCR4 (Fig. 2c). To
determine if the increase in expression relates to an increase in cell surface receptor levels, NHBE ALI cultures were exposed to CS as described in Fig. 2. Following an additional 24 hours,
cell surface CD4 levels were determined by single cell imaging flow cytometry. Cells were immuno-stained with surface markers using anti-human CD4 (BD Biosciences, Cat # 555346,) labeled
with FITC. Isotype controls (BD Biosciences, Cat # 555748) were also analyzed to account for nonspecific staining. Data were acquired and analyzed with Amnis® FlowSight® Imaging Flow
Cytometer and IDEAS software. Staining, image collection, and analysis were carried out according to manufacturer’s protocol and as previously published21,22. As seen in Fig. 3, panels a-d,
smoked NHBE cells showed a significantly increased percentage of CD4+/FITC+ cells (32.78 ± 2.71) compared to air NHBE cells (panel a). Mean fluorescent intensity (MFI) of FITC in the total
population of cell images acquired (10,000 per sample) shows that concurrent with % gated cells, smoked NHBE cells show increased MFI compared to air NHBE cells (panel b). A representative
overlay histogram of intensity of FITC for isotype control (yellow), air (blue) and smoke (green) shows a rightward shift in intensity of smoked cells indicate an increase in CD4 intensity
in smoked NHBE cells compared to air-exposed NHBE cells (panel c). The representative single cell images for each sample clearly shows an increased surface expression of CD4 in smoke exposed
cells compared to air-exposed controls (panel d). To determine if increased expression of CCR5 correlates with increased cell surface expression of CCR5, NHBE ALI cultures exposed to CS
were immuno-stained with surface markers using anti-human CCR5 (BD Biosciences, Cat # 561748,) labeled with APC. Isotype controls (BD Biosciences, Cat # 555576,) were also analyzed to
account for non-specific staining. As seen in Fig. 4, panels a–d, NHBE ALI cultures exposed to CS showed a significantly increased percentage of CCR5+ /APC+ cells (20.25 ± 4.5%, P = 0.03)
compared to air-exposed NHBE ALI cultures (8.58 ± 2.85%) (panel a). MFI of APC in the total population of cell images acquired (10,000 per sample) shows that concurrent with % gated cells,
smoked NHBE cells show increased MFI compared to air NHBE cells (panel b). A representative overlay histogram of intensity of APC for isotype control (yellow), air (blue) and smoke (green)
shows a rightward shift in intensity of smoked cells indicate an increase in CCR5 intensity in smoked NHBE cells compared to air NHBE cells (panel c). The representative single cell images
for each sample clearly shows an increased surface expression of CCR5 in smoke exposed cells compared to air-exposed controls (panel d). We did observe an increased background staining with
our isotype controls for both CD4 and CCR5. However, in both cases, staining was intracellular suggesting a non-specific uptake mechanism of the IgG antibody possibly due to presence of Fc
receptors on NHBE cells. Bronchial epithelium of human, non-human primate, and mouse express Fc receptors that are involved in Fc receptor-dependent transcytosis of IgG across mucosal
barriers23. CS INCREASES CCR5 EXPRESSION BY TGF-Β MEDIATED SUPPRESSION OF MIR-141-5P A number of reports including ours have shown that CS induces TGF-β signaling11,24,25. TGF-β signaling
has also been shown to directly suppress several microRNAs including miR-141-5p26, the only experimentally validated miRNA known to tightly regulate CCR5 expression via ribosomal
frameshifting27. Primarily, we have investigated if increase in CCR5 protein levels is due to an overall increase in CCR5 mRNA. NHBE ALI cultures were exposed to CS (air as control). Total
RNA was isolated and analyzed for CCR5 mRNA levels as described by us earlier5. As seen in Fig. 5a, CS increases CCR5 mRNA levels suggesting that increase in CCR5 by CS is due to increase in
CCR5 transcripts. Given that CS induces TGF-β signaling, we tried to determine if TGF-β treatment leads to a similar increase in CCR5 mRNA levels. NHBE ALI cultures were treated with TGF-β
(vehicle as control) as described by us earlier11. As seen in Fig. 5b, TGF-β treatment also increases CCR5 mRNA levels. Given that TGF-β suppresses miR-141-5p in other cell types, we tried
to determine if TGF-β and CS also suppress miR-141-5p. We analyzed the total RNA for miR-141-5p expression using miR-141-5p specific probe (Life Technologies, Cat # 478712_mir). Figure
5(c,d) shows that both, CS and TGF-β suppress miR-141-5p suggesting that CCR5 induction by CS is via TGF-β-mediated suppression of miR-141-5p. CS AND HIV INDIVIDUALLY AND ADDITIVELY SUPPRESS
CFTR BIOGENESIS AND FUNCTION Given that CS enhances HIV infection of NHBE cells (Fig. 1) and HIV Tat and CS suppress CFTR mRNA (and consequently function) via a common pathway involving
TGF-β signaling5,11, and CS directly suppresses CFTR function by localizing them in aggregosomes28, we tried to determine if HIV infection and CS exposure leads to a synergistic suppression
of CFTR biogenesis and function. We decided on the long-term smoke and HIV exposure to recapitulate the infection process and HIV Tat expression _in vivo_. This is because our observed
kinetics with HIV infection of NHBE ALI cultures demonstrates an initial burst of viral output which gradually decreases and achieves steady state p24 production by day 9 and continues up to
day 505. NHBE ALI cultures were exposed to CS alone as described earlier or CS followed by HIV infection of NHBE ALI cultures. For these studies, we used a lower HIV dose of 2.5 ng/ml p24
equivalent to prevent disruption of the pseudostratified epithelial layer by HIV. Smoke exposed cells (air exposed as control) were infected with either R5 (HIV BaL) or X4 (HIV IIIB) tropic
strain of HIV and exposed to chronic smoke exposure on days 0, 3, 6 and 9 post-infection (air as control). Total RNA was extracted and analyzed for CFTR mRNA levels by qRT-PCR. Exposure to
CS alone or infection with either R5- or X4-tropic HIV by itself suppressed CFTR mRNA (Fig. 6a) and function (Fig. 6b). However, CS exposure exacerbated CFTR mRNA suppression in NHBE ALI
cultures also infected with HIV. Another set of lung-matched NHBE ALI cultures grown on snap wells were treated identically and mounted in Ussing chambers. CFTR activation in response to
β2-agonist albuterol was determined as described by us earlier5,11. DISCUSSION The airway epithelium comprises a primary barrier to occlude inhaled particles such as allergens, toxicants
(CS) and pathogens. Exposure to CS promotes various pathophysiology mechanisms like oxidative stress and activation of CYP enzymes15,16 suppression of CFTR biogenesis11,29, ciliophagy and
consequently MCC in NHBE cells11,30. Several reports have demonstrated an increased susceptibility to lung comorbidities like pneumonia, chronic bronchitis, asthma and COPD in cART era HIV
population compared to non-infected age-matched controls31,32. These effects are exacerbated in smokers and upon cART cessation providing a clear pointer to the role of HIV replication and
smoking in these lung diseases. Rao _et_ _al_. have already demonstrated that oxidative stress due to CS enhances HIV replication15. To date no studies have addressed the interplay between
CS and HIV infection in airway epithelial cells. In this manuscript, we demonstrate that CS also enhances viral entry by upregulating HIV co-receptor CCR5. We observed that CS only enhances
infection of R5-tropic strain of HIV but not X4-tropic strain in age and lung-matched NHBE ALI cultures. This indicates that smoke does not affect stages downstream of viral reverse
transcription. Moreover, when NHBE ALI cultures are infected with the single cycle HIV RGH virus enveloped with R5-tropic envelope, increased levels of proviral DNA are detected in smoke
exposed cells suggesting that the increase in HIV replication is due to enhanced entry by HIV. Given that the NHBE ALI cultures are age- and lung-matched, and increased HIV p24 and RNA is
only observed in R5-tropic HIV infected cells but not X4-tropic HIV infected cells, also rules out the possibility that increase in viral entry is due to changes in non-canonical receptors.
Since the only difference in the life cycle of the two strains is co-receptor usage we tried to determine if CS increases expression of HIV receptors. We found that while CS increases
expressions of CD4 and CCR5, it does not alter expression of CXCR4. Our data also shows that CS mediated increases in protein levels correlate with flow cytometry data of cell surface CCR5
and CD4. Our data also suggests that co-receptor expression on NHBE cells is the limiting factor determining HIV infection. Indeed, NHBE ALI cultures and bronchial brushings from HIV
patients expressed very high levels of CD4 (7-fold compared to CFTR) but lower levels of co-receptor CXCR4 and CCR55. Since CS enhances TGF-β signaling (we have already shown effects of
smoke on CFTR are mediated by TGF-β), we tried to determine if TGF-β plays a role in CCR5 upregulation. We found that TGF-β also upregulates CCR5 expression. Given reports that TGF-β
suppresses several microRNAs including miR-141-5p26, the only experimentally validated miRNA known to regulate CCR5, we tried to determine if TGF-β and by extension CS also suppresses
miR-141-5p in NHBE ALI cultures. miRNAs are endogenous non-coding RNAs of ~22 nucleotides length which mediate their regulatory effects by binding the 3′-UTR binding of the target mRNAs,
causing either cleavage or translational repression based on sequence complementarity. Our data validate that CS and TGF-β both suppress miR-141-5p suggesting that CS upregulates CCR5 via
the TGF-β/miR-141-5p axis. Hence exogenous delivery for miR-141-5p can serve as an important therapeutic lead to suppress CCR5 expression and protect the bronchial epithelium from infection
by R5-tropic HIV. Finally, we tried to determine if the increased interplay between CS and HIV exacerbates CFTR given that CS and HIV Tat suppress CFTR via a common pathway involving TGF-β
signaling5,11. We found that both R5-tropic and X4-tropic strains of HIV and CS individually and additively suppress CFTR mRNA and consequently its function. This could be because CS
enhances viral replication15, thereby increasing the Tat burden. At the same time CS and Tat also activate TGF-β signaling. We observed maximal CFTR suppression in CS exposed NHBE ALI
cultures infected with R5-tropic strain possibly because CS also enhances CCR5 expression thereby increasing viral entry. Hence CS can exacerbate the effects of HIV infection in the airway
with a greater exacerbation observed with R5-tropic strain of HIV. Figure 7 demonstrates a schematic representation of the interplay between CS and HIV infection whereby CS enhances viral
replication and entry (at least in case of R5-tropic virus). CS and Tat suppresses miR-141-5p with a concomitant upregulation of CCR5 co-receptor. Increased infection and replication will
increase the HIV Tat burden in the airway that, with CS additively suppresses CFTR biogenesis and function. While cell to cell transmission is also responsible for HIV infection, due to
limitations in the culture conditions for NHBE ALI cultures (ALI medium does not allow immune cell growth5) we were unable to test effects of CS on cell to cell transmission of HIV. However,
we believe that this would not alter our observations since cell to cell transmission also requires fusion by viral gp120 on infected cells with co-receptors on target cells33. Also cell
free virus has been used as proxy for infection studies with NHBE cultures5,34. In conclusion, our findings suggest that HIV smokers may demonstrate increased CFTR dysfunction compared to
their non-smoking counterparts. Given that CFTR plays a pivotal role in MCC and several other aspects of airway innate immunity, increased CFTR dysfunction in HIV smokers can explain the
increased incidence of lung infections in HIV smokers compared to HIV non-smokers. MATERIALS AND METHODS Primary human bronchial epithelial cells were obtained as described by Fulcher and
Randall35,36 and re-differentiated at the ALI as reported by us11,37. Donor consent was obtained for research if lungs were found unsuitable for transplantation for reasons unrelated to
airway disease and provided by the University of Miami Life Alliance Organ Recovery Agency (LAORA). Since the material is obtained from deceased individuals with minor, de-identified
information, its use does not constitute human subjects research as defined by CFR 46.102. LAORA maintains a signed consent of each individual (or legal healthcare proxy) for donation of the
lungs for research. Experiments used cells from non-smokers to not confound the findings. The primary cultures undergo differentiation at the ALI reproducing the _in vivo_ morphology and
key physiologic processes to regenerate the native bronchial epithelium _ex vivo_35,36. CS EXPOSURE NHBE ALI cultures were exposed to CS using a SCIREQ smoke robot. Four cigarettes were
smoked with a puff volume of 35 ml for 2 seconds every 60 seconds and blown over cell culture filter at rate of 5 ml/min according to ISO 3308. The total number of puffs was 32 for a
duration of approximately 35 minutes. Smoke exposures were done 24 hours prior to infection with R5 or X4-tropic HIV. For chronic smoke exposure, cells were exposed to CS every 3 days using
the same regimen. VIRUS STRAINS AND INFECTION STUDIES The X4-tropic viral strain HIV IIIB was a kind gift from Dr. John Rossi (City of Hope Medical Center, Duarte, CA) and R5-tropic viral
strain HIV BaL was a kind gift from Dr. Madhavan Nair (Department of Immunology, Institute of Neuroimmune Pharmacology, Herbert Wertheim College of Medicine, Florida International
University. Miami, FL). NHBE ALI cultures grown on transwells were infected apically and basolaterally with of either X4-tropic HIV strain (IIIB) or R5-tropic HIV strain (BaL) (5 ng/ml p24
equivalent) as described earlier5. 16 hours post infection, cells were washed apically and basolaterally with PBS four times to remove any residual input virus. The fourth wash was collected
for p24 analysis and measured as day 0 to confirm that all input virus had been removed. Culture supernatants were collected on day 3 and p24 antigen levels were determined using p24 ELISA
kit (ZeptoMetrix Corp. Cat # 0801200) according to manufacturer’s instructions. For chronic HIV exposure, NHBE ALI cultures grown in snap wells cells were infected with X4-tropic HIV strain
(IIIB) or R5-tropic HIV strain (BaL) (2.5 ng/ml p24 equivalent). 16 hours post-infection, cells were washed apically and basolaterally with PBS four times to remove any residual input virus.
Infection was allowed to proceed for 9 days. For single cycle HIV infection, RGH virus was enveloped with R5-tropic envelope as reported by us earlier5. NHBE ALI cultures were infected with
R5-tropic RGH virus. 16 hours post-infection, cells were washed apically and basolaterally with PBS four times to remove any residual input virus. 3 days post-infection, total DNA was
isolated and quantitated for HIV LTR as reported in our earlier manuscript. Copy numbers were determined using comparative Ct method with known copies of HIV DNA. ELECTROPHYSIOLOGY
EXPERIMENTS CFTR activation was determined by Ussing chamber methods as reported by us earlier11. The initial resistance of each filter was measured by application of 1 mV bipolar pulses of
2 seconds’ duration. Amiloride (10 µM) was added apically to inhibit epithelial sodium channel (ENaC) influences. To measure CFTR, albuterol was added and short circuit current (ΔIsc) was
allowed to stabilize. ΔISC in response to albuterol was determined as an index of CFTR function. QUANTITATIVE RT-PCR For qRT-PCR analysis total RNA was extracted using the Qiagen RNeasy mini
kit (Cat # 74104) and complementary DNA (cDNA) was reverse transcribed using the Applied Biosystems high-capacity cDNA reverse transcription kit (Cat # 4368814). qRT-PCR was performed on
the Bio-Rad CFX96 real-time system using validated TaqMan probes (Life Technologies/Applied Biosystems CD4, Cat # HS01058407-m1; CXCR4, Cat # HS00607978-s1;CCR5, Cat # HS99999149-s1; CFTR,
Cat # HS00357011-m1 and GAPDH, Cat # Hs02758991_g1). qRT-PCR results are represented as relative quantification normalized against internal control (GAPDH). MIR-141-5P QPCR EXPERIMENTAL
PROTOCOL NHBE ALI cultures were exposed to CS (air as control) or TGF-β (vehicle as control). Following 24 hours total RNA was extracted from exposed to CS and TGF-β treated with NHBE ALI
cultures by using the Qiagen RNeasy mini kit (Cat # 74104) and cDNA was reverse transcribed by the Applied Biosystems TaqMan™ Advanced miRNA cDNA Synthesis Kit (Life Technologies/Applied
Biosystems, Cat # A28007) according to the manufacturer’s instructions. Real time qPCR was done using TaqMan™ Fast Advanced Master Mix (Life Technologies/Applied Biosystems, Cat # 4444557)
in combination with validated TaqMan probes (Life Technologies/Applied Biosystems, hsa-miR-141-5p, Cat # 478712_mir) according to the manufacturer’s directions. qRT-PCR results are
represented as relative quantification normalized against internal control (GAPDH). WESTERN BLOT ANALYSIS Total protein was loaded onto a gel and run at 100 V. Protein was transferred to
PVDF membrane. Following blocking in 5% milk primary antibodies; CD4 (1: 500; Sigma-Aldrich, Cat # HPA004252), CXCR4 (1:1000; ThermoFisher scientific, Cat # PA5-19857), CCR5 (1:1000;
ThermoFisher scientific, Cat # PA1-41303) and β- actin (1:1000; Cell Signaling, Cat # 4970) were added. Blot was incubated in an anti-rabbit secondary antibody diluted to a concentration of
1:2500. Bands were detected in Chemidoc (Bio-Rad Laboratories, USA) using supersignal west femto maximum sensitivity substrate (ThermoFisher scientific, Cat # 34095) in accordance with the
manufacturer’s instructions. Quantitative densitometry analyses were performed using the Quantity One software (Bio-Rad Laboratories, USA) and the density values are normalized to β-actin.
FLOW CYTOMETRY ANALYSIS NHBE ALI cultures were exposed to CS. At the end of smoking, cells were immuno-stained with surface markers anti-Human CD4 (BD Biosciences, Cat # 555346,) labeled
with FITC, anti-human CCR5 (BD Biosciences, Cat # 561748) labelled with APC. Isotype controls (BD Biosciences, Cat # 555748, Cat # 555576,) were also analyzed to account for nonspecific
staining. Data were acquired with Amnis® FlowSight® Imaging Flow Cytometer. Staining and image collection were carried out according to manufacturer’s protocol and as previously published22.
Images for compensation were collected with compensation beads (552843, BD Biosciences) labeled with the same antibody. STATISTICAL ANALYSIS Unless otherwise mentioned, data were expressed
as mean ± SEM from NHBE ALI cultures from at least three lungs. The data were subjected to statistical analysis using unpaired t-tests for two groups or ANOVA followed by Tukey Kramer
honestly significant difference test for multiple comparisons as appropriate. The significance was considered at the level of P < 0.05. REFERENCES * Dickson, S. J. _et al_. Survival of
HIV-infected patients in the intensive care unit in the era of highly active antiretroviral therapy. _Thorax_ 62, 964-968, doi:thx.2006.072256 [pii] doi:10.1136/thx.2006.072256 (2007). *
Crothers, K. _et al_. HIV infection and risk for incident pulmonary diseases in the combination antiretroviral therapy era. _American journal of respiratory and critical care medicine_ 183,
388–395, https://doi.org/10.1164/rccm.201006-0836OC (2011). Article PubMed PubMed Central Google Scholar * Gordin, F. M. _et al_. Pneumonia in HIV-infected persons: increased risk with
cigarette smoking and treatment interruption. _American journal of respiratory and critical care medicine_ 178, 630–636, https://doi.org/10.1164/rccm.200804-617OC (2008). Article PubMed
PubMed Central Google Scholar * Kohli, R. _et al_. Bacterial pneumonia, HIV therapy, and disease progression among HIV-infected women in the HIV epidemiologic research (HER) study. _Clin
Infect Dis_ 43, 90–98, https://doi.org/10.1086/504871 (2006). Article PubMed Google Scholar * Chinnapaiyan, S. _et al_. HIV Infects Bronchial Epithelium and Suppresses Components of the
Mucociliary Clearance Apparatus. _PloS one_ 12, e0169161, https://doi.org/10.1371/journal.pone.0169161 (2017). Article PubMed PubMed Central CAS Google Scholar * Wanner, A., Salathe, M.
& O’Riordan, T. G. Mucociliary clearance in the airways. _American journal of respiratory and critical care medicine_ 154, 1868–1902 (1996). Article PubMed CAS Google Scholar *
Sethi, S. & Murphy, T. F. Bacterial infection in chronic obstructive pulmonary disease in 2000: a state-of-the-art review. _Clin Microbiol Rev_ 14, 336–363,
https://doi.org/10.1128/CMR.14.2.336-363.2001 (2001). Article PubMed PubMed Central CAS Google Scholar * Hirschtick, R. E. _et al_. Bacterial pneumonia in persons infected with the
human immunodeficiency virus. Pulmonary Complications of HIV Infection Study Group. _The New England journal of medicine_ 333, 845–851, https://doi.org/10.1056/NEJM199509283331305 (1995).
Article PubMed CAS Google Scholar * Fichtenbaum, C. J., Woeltje, K. F. & Powderly, W. G. Serious Pseudomonas aeruginosa infections in patients infected with human immunodeficiency
virus: a case-control study. _Clin Infect Dis_ 19, 417–422 (1994). Article PubMed CAS Google Scholar * Lyczak, J. B., Cannon, C. L. & Pier, G. B. Lung infections associated with
cystic fibrosis. _Clin Microbiol Rev_ 15, 194–222 (2002). Article PubMed PubMed Central CAS Google Scholar * Unwalla, H. J., Ivonnet, P., Dennis, J. S., Conner, G. E. & Salathe, M.
Transforming growth factor-beta1 and cigarette smoke inhibit the ability of beta2-agonists to enhance epithelial permeability. _Am J Respir Cell Mol Biol_ 52, 65–74,
https://doi.org/10.1165/rcmb.2013-0538OC (2015). Article PubMed PubMed Central CAS Google Scholar * Bracke, K. R. _et al_. Cigarette smoke-induced pulmonary inflammation, but not airway
remodelling, is attenuated in chemokine receptor 5-deficient mice. _Clin Exp Allergy_ 37, 1467–1479, https://doi.org/10.1111/j.1365-2222.2007.02808.x (2007). Article PubMed CAS Google
Scholar * Wang, F. & He, B. CCR1 and CCR5 expression on inflammatory cells is related to cigarette smoking and chronic obstructive pulmonary disease severity. _Chin Med J (Engl)_ 125,
4277–4282 (2012). ADS CAS Google Scholar * Sato, K. _et al_. TGF-beta 1 reciprocally controls chemotaxis of human peripheral blood monocyte-derived dendritic cells via chemokine
receptors. _Journal of immunology_ 164, 2285–2295 (2000). Article CAS Google Scholar * Rao, P., Ande, A., Sinha, N., Kumar, A. & Kumar, S. Effects of Cigarette Smoke Condensate on
Oxidative Stress, Apoptotic Cell Death, and HIV Replication in Human Monocytic Cells. _PloS one_ 11, e0155791, https://doi.org/10.1371/journal.pone.0155791 (2016). Article PubMed PubMed
Central CAS Google Scholar * Ande, A., McArthur, C., Kumar, A. & Kumar, S. Tobacco smoking effect on HIV-1 pathogenesis: role of cytochrome P450 isozymes. _Expert Opin Drug Metab
Toxicol_ 9, 1453–1464, https://doi.org/10.1517/17425255.2013.816285 (2013). Article PubMed PubMed Central CAS Google Scholar * Chinnapaiyan, S. & Unwalla, H. J. Mucociliary
dysfunction in HIV and smoked substance abuse. _Front Microbiol_ 6, 1052, https://doi.org/10.3389/fmicb.2015.01052 (2015). Article PubMed PubMed Central Google Scholar * Ande, A. _et
al_. Effect of mild-to-moderate smoking on viral load, cytokines, oxidative stress, and cytochrome P450 enzymes in HIV-infected individuals. _PloS one_ 10, e0122402,
https://doi.org/10.1371/journal.pone.0122402 (2015). Article PubMed PubMed Central CAS Google Scholar * Abbud, R. A., Finegan, C. K., Guay, L. A. & Rich, E. A. Enhanced production
of human immunodeficiency virus type 1 by _in vitro_-infected alveolar macrophages from otherwise healthy cigarette smokers. _The Journal of infectious diseases_ 172, 859–863 (1995). Article
PubMed CAS Google Scholar * Dahabieh, M. S., Ooms, M., Simon, V. & Sadowski, I. A doubly fluorescent HIV-1 reporter shows that the majority of integrated HIV-1 is latent shortly
after infection. _J Virol_ 87, 4716–4727, https://doi.org/10.1128/JVI.03478-12 (2013). Article PubMed PubMed Central CAS Google Scholar * Parira, T. _et al_. Novel detection of
post-translational modifications in human monocyte-derived dendritic cells after chronic alcohol exposure: Role of inflammation regulator H4K12ac. _Scientific reports_ 7, 11236,
https://doi.org/10.1038/s41598-017-11172-6 (2017). Article ADS PubMed PubMed Central CAS Google Scholar * Figueroa, G. _et al_. Characterization of Human Monocyte-derived Dendritic
Cells by Imaging Flow Cytometry: A Comparison between Two Monocyte Isolation Protocols. _J Vis Exp_, https://doi.org/10.3791/54296 (2016). * Spiekermann, G. M. _et al_. Receptor-mediated
immunoglobulin G transport across mucosal barriers in adult life: functional expression of FcRn in the mammalian lung. _J Exp Med_ 196, 303–310 (2002). Article PubMed PubMed Central CAS
Google Scholar * Churg, A., Tai, H., Coulthard, T., Wang, R. & Wright, J. L. Cigarette smoke drives small airway remodeling by induction of growth factors in the airway wall. _American
journal of respiratory and critical care medicin_e 174, 1327–1334, 200605-585OC [pii] doi:10.1164/rccm.200605-585OC (2006). * Wang, R. D., Wright, J. L. & Churg, A. Transforming growth
factor-beta1 drives airway remodeling in cigarette smoke-exposed tracheal explants. _Am J Respir Cell Mol Biol_ 33, 387–393, 2005-0203OC [pii] doi:10.1165/rcmb.2005-0203OC (2005). * Guo, L.
_et al_. MicroRNAs, TGF-beta signaling, and the inflammatory microenvironment in cancer. _Tumour Biol_ 37, 115–125, https://doi.org/10.1007/s13277-015-4374-2 (2016). Article PubMed CAS
Google Scholar * Belew, A. T. _et al_. Ribosomal frameshifting in the CCR5 mRNA is regulated by miRNAs and the NMD pathway. _Nature_ 512, 265–269, https://doi.org/10.1038/nature13429
(2014). Article ADS PubMed PubMed Central CAS Google Scholar * Clunes, L. A. _et al_. Cigarette smoke exposure induces CFTR internalization and insolubility, leading to airway surface
liquid dehydration. _FASEB J_ 26, 533–545, fj.11-192377 [pii] doi:10.1096/fj.11-192377 (2012). * Cloonan, S. M., Lam, H. C., Ryter, S. W. & Choi, A. M. “Ciliophagy”: The consumption of
cilia components by autophagy. _Autophagy_ 10, 532–534, https://doi.org/10.4161/auto.27641 (2014). Article PubMed PubMed Central Google Scholar * Pace, E. _et al_. Comparative
cytoprotective effects of carbocysteine and fluticasone propionate in cigarette smoke extract-stimulated bronchial epithelial cells. _Cell stress & chaperones_ 18, 733–743,
https://doi.org/10.1007/s12192-013-0424-0 (2013). Article CAS Google Scholar * Diaz, P. T. _et al_. Increased susceptibility to pulmonary emphysema among HIV-seropositive smokers. _Ann
Intern Med_ 132, 369–372 (2000). Article PubMed CAS Google Scholar * Morris, A. _et al_. HIV and chronic obstructive pulmonary disease: is it worse and why? _Proceedings of the American
Thoracic Society_ 8, 320–325, https://doi.org/10.1513/pats.201006-045WR (2011). Article PubMed PubMed Central Google Scholar * Puigdomenech, I., Massanella, M., Cabrera, C., Clotet, B.
& Blanco, J. On the steps of cell-to-cell HIV transmission between CD4 T cells. _Retrovirology_ 6, 89, https://doi.org/10.1186/1742-4690-6-89 (2009). Article PubMed PubMed Central CAS
Google Scholar * Brune, K. A. _et al_. HIV Impairs Lung Epithelial Integrity and Enters the Epithelium to Promote Chronic Lung Inflammation. _PloS one_ 11, e0149679,
https://doi.org/10.1371/journal.pone.0149679 (2016). Article PubMed PubMed Central CAS Google Scholar * Fulcher, M. L., Gabriel, S., Burns, K. A., Yankaskas, J. R. & Randell, S. H.
Well-differentiated human airway epithelial cell cultures. _Methods Mol Me_d 107, 183–206, doi:1-59259-861-7:183 [pii] (2005). * Fulcher, M. L. & Randell, S. H. Human nasal and
tracheo-bronchial respiratory epithelial cell culture. _Methods Mol Biol_ 945, 109–121, https://doi.org/10.1007/978-1-62703-125-7_8 (2013). Article PubMed CAS Google Scholar * Unwalla,
H. J., Horvath, G., Roth, F. D., Conner, G. E. & Salathe, M. Albuterol modulates its own transepithelial flux via changes in paracellular permeability. _Am J Respir Cell Mol Biol_ 46,
551–558, https://doi.org/10.1165/rcmb.2011-0220OC (2012). Article PubMed PubMed Central CAS Google Scholar Download references ACKNOWLEDGEMENTS Financial support from National Institute
of Health grant R21-HL128141-01A1. AUTHOR INFORMATION AUTHORS AND AFFILIATIONS * Department of Immunology, Institute of Neuroimmune Pharmacology, Herbert Wertheim College of Medicine,
Florida International University, Miami, FL, USA S. Chinnapaiyan, R. Dutta, J. Bala, T. Parira, M. Agudelo, M. Nair & H. J. Unwalla Authors * S. Chinnapaiyan View author publications You
can also search for this author inPubMed Google Scholar * R. Dutta View author publications You can also search for this author inPubMed Google Scholar * J. Bala View author publications
You can also search for this author inPubMed Google Scholar * T. Parira View author publications You can also search for this author inPubMed Google Scholar * M. Agudelo View author
publications You can also search for this author inPubMed Google Scholar * M. Nair View author publications You can also search for this author inPubMed Google Scholar * H. J. Unwalla View
author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS C.S. prepared the manuscript and performed the experiments. D.R. performed the experiments. B.J.
Formal analysis. P.T. performed the experiments. A.M. analyzed the data. N.M. prepared the manuscript. U.H.J. prepared the manuscript, performed experiments and analyzed the data.
CORRESPONDING AUTHOR Correspondence to H. J. Unwalla. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE:
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. ELECTRONIC SUPPLEMENTARY MATERIAL SUPPLEMENTARY DATA RIGHTS AND
PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any
medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The
images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not
included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly
from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Chinnapaiyan, S.,
Dutta, R., Bala, J. _et al._ Cigarette smoke promotes HIV infection of primary bronchial epithelium and additively suppresses CFTR function. _Sci Rep_ 8, 7984 (2018).
https://doi.org/10.1038/s41598-018-26095-z Download citation * Received: 19 December 2017 * Accepted: 04 May 2018 * Published: 22 May 2018 * DOI: https://doi.org/10.1038/s41598-018-26095-z
SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to
clipboard Provided by the Springer Nature SharedIt content-sharing initiative